Effect of Sensory Substitution Devices on Children
Info: 7256 words (29 pages) Dissertation
Published: 16th Dec 2019
Tagged: Psychology
INTRODUCTION:
Sensory substitution devices are those which allow for the compensation of a lost sense by allowing another sense to convey the same information that would be garnered by the lost sense to the brain. The first sensory substitution devices have been in study, with the first invented being the walking cane and braille (Ojala, 2016). Paul Bach-y-Rita was one of the first to seriously believe that neuroplasticity was still present in adult brains, and developed one of the first sensory substitution systems. His work in the 1960s did much to advance the field, as he introduced a chair that worked to allow the blind to view their surroundings. This device was named TTVS and functioned by using a camera to record what was within the user’s eye range, the stimuli were then translated into an assortment of vibrating tactile stimuli located on the back of the chair where the subject was sitting. In this way, the chair could convey information about shadows, items, and faces present in the room (Bach-y-Rita P., 1969). Bach-y-Rita’s work would help open a new field of study on the effects of neuroplasticity and the extent to which it was present in the adult brain.

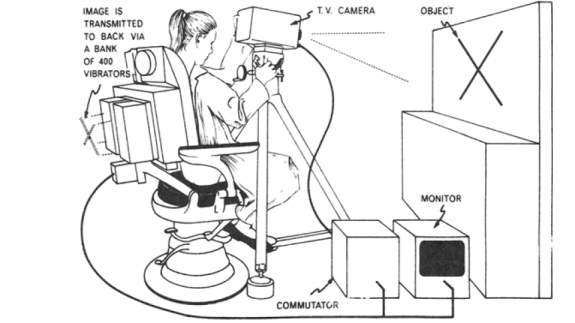
Figure 1: Illustration of Bach-y-Rita’s TTVS. Source: (Benjamin W. White, 1970)
Other sensory substitution devices have been made throughout the years that have made advancement in the translation of visual information to tactile stimuli. In general, sensory substitution devices act by allowing visual information that would normally be detected by the eyes to be processed by another sensory modality. One way in which this may work is to allow for blind individuals to be able to perceive his/her surroundings from the information that is sent from his/her sense of touch to his/her visual cortex. To be clear, the blind person does not directly sense the objects, as a sighted person would, but rather he/she begins to associate the object directly with the touch that signifies its presence—meaning that they perceive the object as being before them, but don’t create a visual image of it in their head. Further, as the perception of seeing the object is created through the sense of touch, it may be possible that the visual cortex is activated using such sensory substitution devices, as well as the somatosensory cortex—as the two areas now form a system for the registration of the visual information being conveyed through tactile stimulation (Ojala, 2016). Thus, brain plasticity plays a vital role in the processes involved in sensory substitution devices, as it allows for the brain to be rewired in a way that allows for these two areas to become intertwined and more involved with each other than would have before been possible. It is with this prolonged cooperation between these two areas that this continuous perception of vision is possible for blind individuals with the use of a sensory substitution device.
The BrainPort V100, is a modern sensory substitution device which translates visual stimuli into pulses applied to the tongue. It was approved was approved by the Food and Drug Administration in 2015, and substitutes vision with tactile stimuli. The sensory substitution device is composed of a 1.5 cm camera, which is found in a discreet pair of sunglasses. It also includes a base unit, which is held by the user and is about the size of a phone. This base unit holds controls for shock intensity, zoom, and light settings. It also houses a central processing unit (CPU) that works to translate the digital image taken by the camera into an assortment of pulses of electricity. The CPU then transmits its data to an electrode array, which measures approximately 9 cm2, and is placed on the user’s tongue. Each electrode on the tongue conveys information from a set of pixels. With the use of this device blind users can distinguish what is within their eye-line (Kendrick, 2009). In effect, this allows them to see, as the electrical pulses delivered to the tongue are conveyed to the brain, which then receives these electoral impulses similarly to how it would receive them from the eyes.
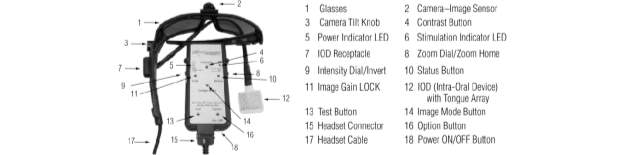
Figure 2: Depiction of the BrainPort device hardware. Source: (Amy C. Nau, 2015)
It has been shown in previous studies that portions of the brain traditionally involved in the interpretation of visual stimuli are recruited for the interpretation of stimuli from other senses when blind subjects undergo training with sensory substitution devices. Such observations have been made in the activation of the occipital cortex (Tomas Ortiz, 2011), as well as the extrastriate body area of the visual cortex (Amedi, 2014), and the lateral-occipital tactile-visual area (Amir Amedi, 2007). It is therefore seen that the brain interprets sensory information in the correct context no matter what sense is garnering the stimulation—so long as it receives the necessary information in the correct format. Thus, although the subject is receiving tactile stimulation, s/he does not attribute the visual information that results to the tactile stimulation, rather s/he attributes it as being external to herself/himself (Kercel, 2003). Thus, in the use of the BrainPort, although the stimulation is received by mechanoreceptors on the tongue, it may be interpreted similarly to the interpretation of visual stimuli, as the subject becomes accustomed to the device and attributes the images s/he sees to belonging in space, rather than on their skin (Kercel, 2003).
The experiment outlined in the remainder of this paper will set out to test multiple thesis’. First, it will test the idea that if the brains of child subjects receive an overall higher percentage of change after training with the BrainPort device, then children, whether sighted of blind, harbor more brain plasticity than adults. This is evidenced by previous studies which have shown that plasticity of the brain is greater at younger ages, because of an excess of neurons and synoptic connections (Johnston MV, 2001). This would lead to the conclusion that blind children should be taught how to use sensory substitution devices at an earlier age than currently accustomed to. This experiment will also test the notion that if there is a larger percentage of change in brain activity in the congenitally blind subjects versus those subjects who lost their vision later in life, when both groups were trained, then plasticity in the brain was greater when no prior use of the visual cortex for seeing was present. This would illustrate that the congenitally blind, who never used their visual cortex for the processing of actual visual stimuli presented more plasticity because their brain didn’t have to break-down any pre-conceived notions about how the visual cortex should function, and was therefore freer to recruit the visual cortex for the processing of tactile stimuli, as received from the BrainPort. This is reinforced by previous work that illustrates the hypercompensation witnessed in subjects who are congenitally or early blind are greated, when compared to other subjects who lost their sight late in life, the performance of which resembles more closely that of sighted individuals (Rieser, 2008). Thus, this would suggest that there are differences in the ability that different populations of blind individuals have for neuroplasticity, a topic that will be tested in this paper. In addition, I have set out to test the hypothesis that if subjects are trained with the BrainPort, the brain plasticity that this evokes will then cause an overall rise in skill when applied to other touch sensitivity measurements—such as vibrotactile detection and spatial gap detection, by lowering the thresholds necessary for both sensations (Add a source?). Thus, I have set out with three propositions to test through experimentation with the use of the BrainPort device.
To test my hypothesis’, I will have twelve groups total. There will be four groups of blind children. Two of these groups will be comprised of children who are congenitally blind with one such group being trained with the BrainPort, while the other will not be trained with the BrainPort. The other two groups will be comprised of children who have lost their sight later in life (from birth to 2 years of age), with one group being trained with the BrainPort and the other group not being trained. Furthermore, there will be four groups of young adults from the ages of 25-30. Two of these groups will be made up of congenitally blind individuals, with one group receiving training with the BrainPort and the other not receiving training. The following two groups of young adults will be made up of individuals who have lost their sight later in their life (20-25 years of age). One of these groups will be trained with the BrainPort, while the other will not receive any training. Included in the study, there will also be four control groups. Two of these groups will be comprised of sighted children, one of which will be trained with the BrainPort while the other will not receive training. The remaining two control groups will be made up of sighted young adults, one of which will train with the BrainPort while the other will
| Congenital blindness | Later on-set blindness | Sighted | Children | Young Adult | Trained | Not trained | |
| Group 1 | X | X | X | ||||
| Group 2 | X | X | X | ||||
| Group 3 | X | X | X | ||||
| Group 4 | X | X | X | ||||
| Group 5 | X | X | X | ||||
| Group 6 | X | X | X | ||||
| Group 7 | X | X | X | ||||
| Group 8 | X | X | X | ||||
|
|
X | X | X | ||||
| Group 10 | X | X | X | ||||
| Group 11 | X | X | X | ||||
| Group 12 | X | X | X | ||||
 Group 9
Group 9The reasoning behind having children among the subjects tested in my research will be to test my hypothesis that the brains ability to train and become accustomed to the BrainPort will be greater in youth. This is reasoned from evidence that shows that children go through a critical period in the establishment of binocular vision from the ages of 1-3 (Martin S. Banks, 1975). In addition, it has been seen that there are several other critical periods in the visual systems, with more specialized layers developing these cemented properties later than others. Overall, the critical periods for vision occur from the opening of the eyes until the puberty (Daw, 1998). These findings illustrate the development of cortical attributes early in life, creating a strong sense for plasticity in the brain of youths. Further, this gives added support for the thought that the children in involved in this experiment will display a heightened mastery of the BrainPort device than their adult counterparts, as their cortical properties will be more malleable to change, leading to them becoming better adept at using sensory substitution devices.
Table 1: Groups and demographic information
Thus, aspects of vision are established in the visual cortex early on in life and continue to govern the interpretation of visual stimuli as an individual matures. Lack of stimuli during these sensitive periods may lead to a deficiency in the eye sight of the child which lasts a life time. This is something that has been seen in the study of cats, in which it has been shown that after a specific period the degrading effects caused by the lack of visual stimuli stop being reversible due to the passing of critical periods (D.H. Hubel, 1970). This is has also been seen in the study of strabismus, when developed early in a child’s life, in which early intervention before the passing of the critical period involved is key to restoring eyesight in both eyes and allowing for the development of binocular vision (Martin S. Banks, 1975). Thus, it can be observed that early intervention is often vital in the development of the mechanisms involved in the interpretation of visual stimuli. It may be that in blind children the same type of early intervention is necessary in order to allow for the sensory substitution device to become fully integrated with the same mechanisms involved in visual interpretation.
The young subjects were selected to be 3 years of age to ensure that none of the critical periods related to the visual system had passed. Thus, I have decided to have the child subjects be as young as possible, while still being able to actively participate in the study.
Furthermore, I will also be using groups made up of individuals with both congenital and later on-set blindness. This will be to allow the comparison of the percentage of differences in brain plasticity between the two groups training with the BrainPort device. In blind individuals, the visual cortex is frequently recruited for use by the other senses this however, is often of greater advantage when vision is lost at birth or early on in life. Whereas, if a person becomes blind later in life, their performance in tactile tasks are much closer in skill to those of their sighted counterparts (Ella Striem-Amit, 2012). This may be due to the brain in individuals who lose their sight later in life having already become adjusted to interpreting visual stimuli received from the retinas in the visual cortex—leaving no room for the said cortex to be recruited by the other senses. Thus, after the loss of vision these patterns of having visual stimuli be interpreted in the visual cortex must first be repressed for the visual cortex to begin to translate tactile stimuli as visual perception—potentially making the process of getting accustomed to sensory substitution devices more challenging and time-consuming. Thus, the other senses are not able to become enhanced with the absence of vision during the same span of time among all blind subjects. In addition, the changes that must be made in the brain to allow for the interpretation of tactile stimuli to visual sensation are made harder because the critical periods in the visual cortex that have been discussed earlier have already passed (Daw, 1998). Although some plasticity remains in the brain, it is not present to the extent that will allow monumental changes in the hard-wiring of the mechanisms in the visual cortex, as would be possible before the passing of these critical periods.
Moreover, the reasoning behind having groups of individuals who have congenital blindness and those who have lost their sight later in life, is to not only allow for the comparison of plasticity in the brain between conditions, as mentioned before, but also to allow for comparison of brain plasticity across age groups. This will be possible as there will be children who have been congenitally blind as well as those who have lost their vision later in life. Thus, we will be able to observe if having visual input early on in life will have an inhibitory effect on the plasticity of the brain, even when the critical period associated with the visual cortex has not yet passed, as compared to those subjects who were born blind and had no such visual input.
The presence of sighted individuals in the study will act as a control group from which to compare the changes that take place in all eight groups of blind individuals. These control groups will allow us to compare the brain plasticity that is present in both sighted children and young adults and to see how this compares to their blind counterparts. This comparison will further enhance our view by allowing us to see if having long-term, continual visual stimuli input in the visual cortex will have a detrimental effect on brain plasticity regarding the allowed changes in the structure of the brain after training with the BrainPort.
The percentage of change among subjects will be measured with the use of fMRI scans. Functional magnetic resonance imaging will be used to avoid the use of radiation on subjects and allow for the repeated examination of the activity in the brain throughout the same session, if necessary, with little to no risk (Watson, 2008). A BrainPort device compatible with the use of an fMRI machine will be used by the subject during the scanning process, while they complete an assortment of shape differentiation tasks. Said shape differentiation tasks will be described in detail within the methods section of this paper. The activity of their brain will then be recorded during the fMRI scanning process. These measurements will be taken before and after training with the BrainPort device and will be converted to percentages for each group. Thus, these percentages will be compared among groups as a measurement of relative brain plasticity present among each demographic in each group tested. In addition, two other tactile sensitivity measurements will be used to rate the skill acquired after training with the BrainPort. To accomplish this, a simple checklist will be introduced which accounts for the lowering of thresholds involved. Thus, the skill that the subject had port-training will be measured relative to the skill that the subject started out with prior to the training. The results will measure the relative amount of improvement at the end of the training with the BrainPort. To make such measurements, each subject will participate in the tactile perception tests that will be described in the remainder of this paper both before and after training with the BrainPort device.
Two tactile perception measurements for touch will be used to test whether generalized learning due to brain plasticity occurs throughout the sense used for sensory substitution—or if such learning is isolated to specific channels. The detection of vibrotactile thresholds will be the first test used. This test will be used to determine which receptors, if any, improve in their performance after training with the BrainPort. By evaluating the extent to which the threshold of each receptor channel decreases, I will be able to determine which receptors are mainly involved in using the BrainPort. This study will also show what receptors are mainly involved in plasticity, as the results of the fMRI scanning will be compared in conjunction with the results garnered from this test. Different contactor sizes and frequencies will be used to test each receptor channel. A large 2.9 cm2 contactor will be used with 250Hz vibration to stimulate the PC channel. The PC channel is optimally responsive to high-frequency vibrations, due to its ability for spatial and temporal summation as well as the large receptive fields of its Pacinian-corpuscle fibers. Further, to stimulate the SA2 channel, a smaller .008 cm2 contactor will be used with a 250Hz frequency. The SAII channel is most responsive to the skin being stretched, making it vital in an individual’s awareness to hand position. This is attributable to the channel having slowly adapting receptors that are sparingly interspersed on the skin in combination with nerve fibers with sizeable receptive fields. In addition, because the size of a contactor doesn’t play a role in the vibrotactile detection thresholds of smaller frequencies, a large 2.9 cm2 contactor will be used to stimulate RA receptors with a 10Hz frequency and SA1 receptors with a 1Hz frequency. The RA channel is optimally responsive to tactile stimulus that are mobile over the skin, in contrast to those which are static. This is a result of the RA channel having fibers that rapidly adapt with minute receptive fields. The SA I channel is optimal in the perception of changes in the three-dimensional characteristics of stimuli, a property that is especially well suited for the reading of Braille. This is attributable to the channel having fibers that slowly adapt and have minute receptive fields (George A. Gescheider J. H., 2010).
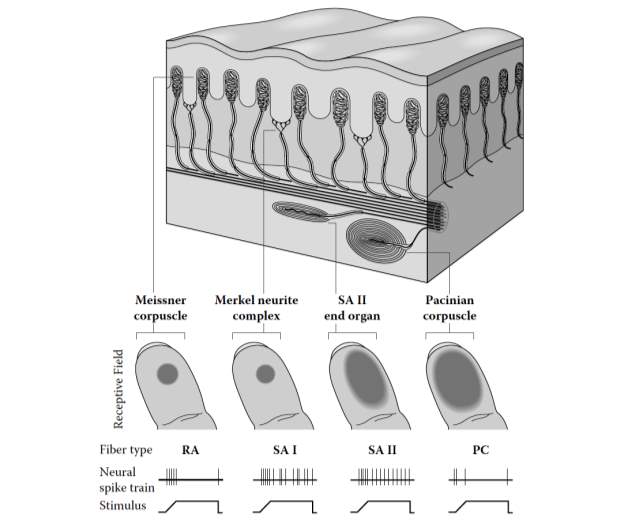
Figure 3: Illustration of nerve fibers and tactile receptors, as found in glabrous skin. Source: (George A. Gescheider J. H., 2010) (George A. Gescheider J. H., 2010)
Moreover, a spatial gap detection task will also be used to measure the relative increase in tactile function after training with the BrainPort. This task will consist of measuring the gap necessary in the stimuli presented for subjects to be able to detect whether there is a gap in said stimuli (ADD Source and further explanation after you read Stevenson article). Thus, the question being asked will be: how big must a gap be for a person to detect said gap? Through this test, I will be able to determine whether detection thresholds see a marked decrease after training with the BrainPort and how this sees variance among age groups and individuals of different conditions. These results will also be compared to those gathered from the fMRI scans to measure the amount of change in plasticity that allows for learning throughout the channels involved in the sense of touch.
METHOD:
There will be three hypotheses that will be tested throughout this research. The first thesis will be the thought that if brains of children exhibit more changes due to training with the BrainPort device, then younger brains harbor more plasticity than those of adults. I will also be testing the idea that if those who are congenitally blind display larger changes in the structure of the brain then those subjects who are congenitally blind will have more brain plasticity that those who are sighted or who have lost their vision later in life. Lastly, I will also set off the test whether the brain plasticity evoked from training with the BrainPort device will also cause an overall rise in skill regarding other measurements of touch sensitivity, this will also allow me to identify which tactile channels are most involved in the changes that occur in the brain.
PARTICIPANTS:
Participants included 12 congenitally blind children (6 received training, while 6 did not). Also, included where 12 children who had lost their sight later in life and within the time span of birth to 2 years of age (6 received training, while 6 did not). Participants also included 12 congenitally blind young adults (6 received training, while 6 did not) and 12 adults who had lost their sight from the ages of 20-25 years old (6 received training, while 6 did not). Control groups were also included in this study to ensure that all observations could be attributed to the etiology behind what caused subjects to lose their sight. Control groups in this study included 12 sighted children (6 received training, while 6 did not) and 12 sighted young adults (6 received training, while 6 did not). In addition, participants were in equal proportions of male to female within each group. These subjects were placed into 12 groups, as can be seen in Table 1. All blind subjects were recruited from the National Federation of the Blind for participation in the study. All participants gave informed written consent and were made aware of all aspects of the study. Subjects were disqualified from the study if they were subject to any impairment, such as brain trauma, that would hamper their ability to utilize the BrainPort device.
INSTRUMENTS:
To effectively test my hypotheses, various tactile detection tasks will be used. The first involves the identification of shapes with the use of the BrainPort device in all subjects. Sighted subjects were blindfolded during the tasks, to ensure that all subjects received equal amounts of external stimuli. The object was displayed on an 18-inch computer screen. This computer was placed on a tabletop, which was covered by a black overlay, and faced a wall with a similar coloration to provide a high contrast environment. Each shape was presented in white on a black background and was placed directly in front of the subject at a distance of 50 cm. The shapes identified included the following: an ellipse, a circle, a square, parallelogram, a triangle, rectangle, a star, and the letters E, A, R, I, O, T, N, S, L, C, U, D, P. The letters were selected due to their high prevalence in use among English speakers (Press, n.d.). The letters were presented with the use of 95-point Times New Roman font. These 20 shapes were presented in a randomized order for each of 10 trials for each subject. This shape differentiation test was completed both before the subjects received any training with the BrainPort and after the subjects received training with the BrainPort. While the subjects completed the task they were being scanned by the fMRI machine and were wearing a BrainPort device that was compatible with the machine. Each subject was given 3 minutes to identify the shape, if no correct response was spoken in this time frame, then the trial was labeled as incorrect. Additionally, each subject was only allowed to answer once, to eliminate broad guesses. Once the answers were gathered from the shape differentiation task, the amount that each subject had named correctly and the time that they spent in giving each correct answer was noted.
After each subject received training with the BrainPort, this shape differenciation task was repeated. The results of the fMRI scan as well as the changes in the amount of correct answers and the time taken to give them were considered for each subject in comparison to the subject’s performance in the same task prior to receiving training. The relationship between these two sets of results were used to translate responses into percentages of change in brain structure to easily evaluate the amount of change in the structure of the brain due to training with the BrainPort. In effect, this will allow us to observe how much brain plasticity there is present in each group tested as a result of acquired skill with the sensory substitution device (Teija Kujala, 2000) (Ron Kupers, 2004).
Additionally, each subject will also participate in two touch sensitivity measurements. These will be used to evaluate if the rewiring of the brain involved in training with the BrainPort device is specialized to one tactile channel or is generalized throughout all tactile channels. The two sensitivity measurements will be the measurement of thresholds in vibrotactile detection and special gap detection.
The vibrotactile detection threshold measurement will be used to determine which receptors, if any, improve in performance after training with the BrainPort device. It is known that the sense of touch is composed of four distinct channels, which remain independent until they converge in the somatosensory cortex (George A. Gescheider J. H., 2012). A single type of mechanoreceptor and afferent nerve fiber is linked to each channel in the tactile sensory system. These channels include the RA channel, composed of Meissner capsules and rapidly adapting (RA) nerve fibers, the PC channel, composed of Parcinian corpuscles and PC nerve fibers, the SA II channel, composed of SA II end organs and slowly adapting Type II, or SA II, nerve fibers, and the SA I channel with its Merkel-neurite complexes and slowly adapting Type I, or SA I, nerve fibers. This specificity in the composition of each tactile channel allows for each to have a high frequency-selectivity. This in turn leads to the possibility that each tactile channel may be innervated and isolated with the careful choosing of intensity and frequency used to test subjects in specific test sites (George A. Gescheider J. H., 2012). Thus, I will identify which receptors are mainly involved in brain plasticity, as the results yielded from this experiment will be compared to those from the fMRI scans, which show changes in activity in the brain. These measurements will also be used to determine which receptors are trained and improved due to training with the BrainPort.
This was be done by utilizing specific frequencies and contactor sizes known to activate each channel. As noted earlier, a large 2.9 cm2 contactor will be used with 250Hz vibration to stimulate the PC channel. A smaller .008 cm2 contactor will be used with a 250Hz frequency to stimulate the SA II channel. In addition, a large 2.9 cm2 contactor will be used to stimulate RA receptors with a 10Hz frequency and SA1 receptors with a 1Hz frequency. The results will be gathered with innervation to the tactile channels located in the thenar eminence on the palm of the hand. The detection thresholds measured from each subject were then plotted as a function of the frequency of the stimulus applied; the resulting graph was then evaluated in comparison to the similarly plotted function as found in previous studies, and as showcased below (George A. Gescheider J. H., 2010).
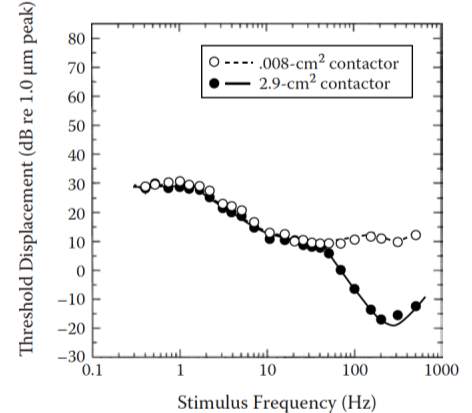
Figure 4: Detection thresholds plotted as function of the applied stimuli, as done via .008 cm2 and 2.9 cm2 contactors. Source: (George A. Gescheider J. H., 2010)
The four neural systems involved in touch facilitate the detection of vibratory stimuli on glabrous skin. Thus, each part of the above function is directly related to a specific tactile channel. This allows for the direct identification of the tactile channel involved when a portion of the curve changes. Therefore, the lowering thresholds and the effect that learning has on the channels can be directly evaluated. This said, the PC fibers determine the thresholds for stimuli with a frequency above 40 Hz, with the use of a large contactor. The SA II fibers dictate the threshold when the stimuli’s frequency is above 100Hz, with the use of a small contactor. The SA I fibers dictate the threshold when the frequency of the stimuli if between 0.4 Hz and 1.5 Hz, regardless of the size of the contactor used. The RA fibers control the thresholds when the frequency of the stimuli is between 1.5 Hz and 40 Hz with the use of a large contactor and those frequencies that are between 1.5 Hz and 100 Hz with the use of a small contactor (George A. Gescheider J. H., 2010).
The detection threshold for each subject was found with the use of an apparatus consisting of a circular contactor shaped to fit the curves of the skins surface. This contactor was bordered by a rigid surround with a 1mm gap present between the two and was attached to a vibrating component of the apparatus. The function of this rigid surround was to limit the extent to which the stimulus could spread throughout the surface of the skin, thereby restraining it to the area directly around the contactor (George A. Gescheider J. H., 2012). The skin temperature was kept constant by a water bath attached to the contactor, which acted to circulate water throughout the apparatus’ hollow chambers. The temperature that the apparatus was kept at was 30°C. This was implemented to reduce the role that skin temperature played in tactile sensitivity (G. A. Gescheider, 1997). Each subject was isolated from all superfluous stimuli with the use of earphones, which delivered narrow-band noise to conceal the sound made by the vibrator, and were placed in a booth that blocked out nonessential external vibration and noise.
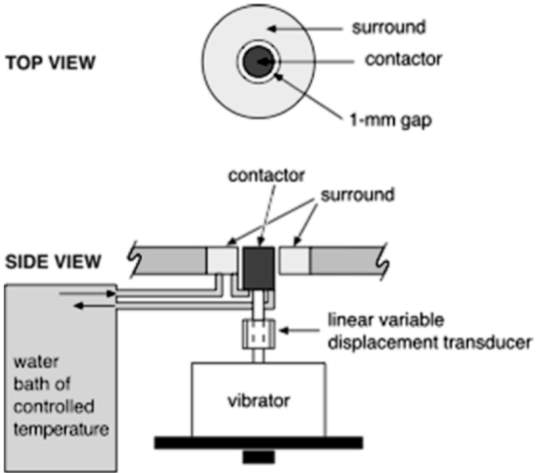
Figure 5: The apparatus used in the measurement of detection thresholds. Source: (George A. Gescheider J. H., 2012)
Moreover, the detection threshold was measured by a single vibratory stimulus, which stimulated either the RA channel, with a frequency of 10 Hz, or the PC channel, with a frequency of 250 Hz. These were issued on the thenar eminence with a 2.9 cm2 contactor. The subjects responded with the use of a two-alternative forced-choice tracking method. The stimulus intensity that led to the subject being correct 75% of the time when correctly detecting the frequency of the presented stimulus, these responses included nonconsecutive correct answers (Jozef J. Zwislocki, 2001). Once the subject’s performance reached to the point that his or her responses were correct 75% of the time, the stimulus intensity was recorded.
These measurements were taken for each subject both before training with the BrainPort and after training with the BrainPort.
The detection thresholds of each subject were measured for a variety of frequencies, to stimulate each tactile channel involved in the processing of tactile stimuli. The detection thresholds gathered from this testing was then plotted as a function of stimulus frequency. The graphs that resulted prior-to and after training were then compared to evaluate which thresholds, if any were lowered, and the tactile channels involved with each of these thresholds. It was with these measurements that we were enabled to conclude whether the rise in skill due to training with the BrainPort was isolated to specific channels or whether the learning was spread throughout all channels. Thus, I was able to conclude which channel, if any in particular, was responsible for the changes seen in the fMRI scans of each subject. This will also allow for the identification of those channels involved in the restructuring of the brain, as the results garnered from the vibrotactile detection thresholds are evaluated in conjunction with the change seen in the fMRI scans as well as that which seen in the gap detection thresholds.
TRAINING WITH THE OPTACON:
All subjects in groups 1, 3, 5, 7, 9 and 11 received training with the BrainPort device. This can be seen in Table 1. These subjects received multi-faceted training. Training included an introduction to the BrainPort and its components, such as the development of familiarization with the device, how to care for it, and how it works to translates stimuli from its visual to its tactile form. The lack of color and depth perception when the device was used was also discussed to give subjects an indication of the realistic limitations of the BrainPort. We then followed the training program as outlined by established studies to maximize the ability of those subjects who were trained with the BrainPort to build their comfortability and familiarity with the device (Amy C. Nau, 2015). Thus, subjects who were trained to use the BrainPort device developed familiarity in recognizing symbols, such as letters and numbers. They were also able to develop tongue-to-hand coordination, the equivalent to hand-eye-coordination in this scenario, and developed skills in navigation while wearing the BrainPort device. In total, the training for subjects took 6 months, during which the subjects participated in at least two 3-hour supervised training sessions twice per day for 3 days each week. This combined to 72 days during the 6-month period in which the subjects trained completed 432 hours of training. Those in groups 2,4,6,8,10, and 12 were not trained and continued with their usual lives during the six-month period.
Each subject will undergo a functional MRI scan four times. This will occur once at the onset of the research, before any subject received training, once two months after the onset of training, once four months after the onset of training, and again at the end of the six-month period of training. Subjects in groups 2,4,6,8,10, and 12, who will not receive any training will also undergo fMRI scans to ensure that all measurements taken are constant among all subjects. During each fMRI scan 20 horizontal slices of the brain will be imaged concurrently to gain a picture of the brain activity throughout the entire brain and to see what parts of the brain are activated due to use of the BrainPort device.
For this study, the parts of the brain that were activated while the BrainPort was in use during the fMRI scan was noted regarding the location of activation in the brain and the percentage of the brain that was activated. This process was repeated for each fMRI scan and the changes among each scan were noted, as the areas of the brain activated while the BrainPort was in use during the fMRI scan were expected to change. These measurements were considered as a measure of brain plasticity, as the structured of the brain involved in the interpretation of tactile stimuli into visual information were altered, as each subject trained with the BrainPort became more accustomed to the device and how it functioned to convey information.
After the six months of training the touch sensitivities of all subjects were once again tested. This included the evaluation and recording of their vibrotactile detection thresholds and gap detection thresholds. The measurements taken from the fMRI scans were converted to percentages of change in brain activation—regarding both the location and extent—to compare the change in each subject’s brain plasticity after training with the BrainPort. The thresholds yielded from each of the two touch sensitivity tests were also recorded, the values measured before training and after training were compared to evaluate the refinement of skill achieved. The vibrotactile detection thresholds were further used to evaluate which channels were trained with the use of the BrainPort device, to evaluate their connection to plasticity in the brain.
The second tactile sensitivity measurement will be taken via a special gap detection test (get Stevenson article and talk about it here)
Works Cited
Amedi, E. S.-A. (2014, March 17). Visual Cortex Extrastriate Body-Selective Area Activation in Congenitally Blind People “Seeing” by Using Sounds. Current Biology, 24(6), 687-692.
Amir Amedi, W. M.-L. (2007, June 01). Shape conveyed by visual-to-auditory sensory substitution activates the lateral occipital complex. Nature Neuroscience, 10(6), 687-689.
Amy C. Nau, C. P. (2015, January 01). Acquisition of Visual Perception in Blind Adults Using the BrainPort Artificial Vision Device. The American Journal of Occupational Therapy, 69(1), 6901290010p1-6901290010p8.
Bach, M. (1994). The Freiburg Visual Acuity Test–Automatic Measurements of Visual Acuity. Optometry and Vision Science, 73(1), 49-53.
Bach-y-Rita P., C. C. (1969, March 8). Vision Substitution by Tactile Image Projection. Nature, 221, 963-964.
Benjamin W. White, F. A.-Y.-R. (1970, January 01). Seeing with the Skin. Perception & Psychophysics, 7(1), 23-27.
D.H. Hubel, T. W. (1970, Feburary). The period of susceptibility to the physiological effects of unilateral eye closure in kittens. The Journal of Physiology, 206(2), 419-436.
Daw, N. W. (1998, April). Critical Periods and Amlyopia. Arch Ophthalmol, 116, 502-505.
Ella Striem-Amit, A. B. (2012). Neurophysiological Mechanisms Underlying Plastic Changes and Rehabilitation following Sensory Loss in Blindness and Deafness. In A. B. Ella Striem-Amit, The Neural Bases of Multisensory Processes (pp. 402-404). Boca Raton, Florida: Taylor & Francis Group.
G. A. Gescheider, J. M. (1997). The effects of skin temperature on the detection and discrimination of tactile stimulation. Somatosensory & motor research, 14(3), 181-188.
George A. Gescheider, J. H. (2010). Information-Processing Channels in the Tactile Sensory System: A Psychophysical and Physiological Analysis. New York, New York: Psychology Press Taylor & Francis Group.
George A. Gescheider, J. H. (2012). Learning in Tactile Channels. Journal of Experiemental Psychology, 38(2), 302-313.
Johnston MV, N. A. (2001). Sculpting the developing brain. Advances in pediatrics, 48, 1-38.
Kendrick, M. (2009, August 13). Tasting the Light: Device Lets the Blind “See” with Their Tongues. Retrieved August 7, 2017, from Scientific American: https://www.scientificamerican.com/article/device-lets-blind-see-with-tongues/
Kercel, P. B.-y.-R. (2003, December). Sensory substitution and the human–machine interface. Trends in Cognitive Science, 7(12), 541-546.
Kilian Schulze-Bonsel, N. F. (2006, March). Visual Acuities “Hand Motion” and “Counting Fingers” Can Be Quantified with the Freiburg Visual Acuity Test. Investigative Ophthalmology and Visual Sciences, 47(3), 1236-1240.
Martin S. Banks, R. N. (1975). Sensitive Period for the Development of Human Binocular Vision. Science, 190, 675-677. Retrieved from http://www.jstor.org/stable/1741203
Ojala, J. (2016, November 30). Sensory Substitution: An Augmented Human Technologies Seminar Paper. Retrieved August 9, 2017, from LinkedIn: https://www.linkedin.com/pulse/sensory-substitution-augmented-human-technologies-seminar-jouni-ojala
Press, O. U. (n.d.). Which letters in the alphabet are used most often? Retrieved August 16, 2017, from English: Oxford Living Dictionaries: https://en.oxforddictionaries.com/explore/which-letters-are-used-most
Rieser, J. (2008). Blindness and brain plasticity in navigation and object perception. New York, NY: Taylor & Francis Group, LLC.
Ron Kupers, M. P. (2004). “Seeing” through the tongue: cross-modal plasticity in the congenitally blind. International Congress Series, 1270, 79-84.
Teija Kujala, K. A. (2000, January 01). Cross-modal reorganization of human cortical functions. Trends in neurosciences (Regular ed.), 23(3), 115-120.
Tomas Ortiz, J. P.-T.-L. (2011, August). Recruitment of Occipital Cortex during Sensory Substitution Training Linked to Subjective Experience of Seeing in People with Blindness. PLos ONE, 6(8), 1-11.
Watson, S. (2008, October 1). How fMRI Works. Retrieved August 12, 2017, from HowStuffWorks.com: http://science.howstuffworks.com/fmri.htm
Cite This Work
To export a reference to this article please select a referencing stye below:
Related Services
View all


Related Content
All TagsContent relating to: "Psychology"
Psychology is the study of human behaviour and the mind, taking into account external factors, experiences, social influences and other factors. Psychologists set out to understand the mind of humans, exploring how different factors can contribute to behaviour, thoughts, and feelings.
Related Articles

DMCA / Removal Request
If you are the original writer of this dissertation and no longer wish to have your work published on the UKDiss.com website then please: