Leptospira: Pathogenesis, Genomics, Antigen, Immune Response and Vaccines
Info: 7467 words (30 pages) Dissertation
Published: 18th Feb 2022
Tagged: BiologyPharmacology
Abstract
Leptospirosis is a zoonotic disease with global occurrence. This review aims in understanding of genomics, pathogenesis, immune response, vaccine design strategies of the disease. the organism has double circular chromosomes, CI (approximately 4.3 Mb) and CII (approximately 350 kb). These are relatively larger than other spirochetes. Leptospires enter the body through small cuts or abrasions, via mucous membranes such as the conjunctiva or through wet skin. Lipopolyshaccride is the main virulent factor of these organisms. Disease due to infection with leptospirosis results in various symptoms, including vasculitis, acute renal failure, jaundice, thrombocytopenia, pulmonary hemorrhage, myocarditis, conjunctival suffusion, and uveitis. Immunity in leptospirosis is predominantly humoral mediated in humans and most animal species. Complement is a key component of the innate immune system responsible for protection against pathogenic leptospiras. There is currently no widely used vaccine for leptospirosis. The first vaccine introduced for leptospirosis was a killed whole cell vaccine that consisted of formalin-killed leptospires. Currently designing techniques for leptospirosis includes bacterines, live attenuated vaccines, and recombinant proteins. advanced molecular studies of the the organism a a timely action which may lead to a novel findings and in designing effective vaccines with a limited side effects and low cost.
Key words: genomics, immunity, leptospira, leptospirosis, pathogenesis, vaccines
Table of Contents
1. Introduction
2. Leptospira at Genomics Level
3. Pathogenesis
3.1. Pathogen Entry and invasion
3.2. Virulence factors
3.2.1. Lipopolysaccharide (LPS)
3.2.2. Loa 22
3.2.3. LipL32
3.2.4. LenA
3.2.5. Hemolysins
3.3. Adhesion and persistence of Leptospira spp. in the host
3.4. Mechanisms of Damage to Host Tissues
4. Antigen and Immune Response to Leptospira infection
4.1. Host Detection of Pathogen
4.1.1. Pattern Recognition Receptors
4.2. Innate Immunity against leptospiral infections
4.2.1. Cellular Response
4.3. Acquired Immunity
4.3.1. Humoral Mediated
4.3.2. Cell-Mediated Immunity
5. Vaccines against Leptospira
5.1. Vaccine design strategies for Leptospira
5.1.1. Bacterin vaccines
5.1.2. Live Vaccines
5.1.3. Lipopolysaccharide Vaccines
5.1.4. Recombinant Proteins as Vaccines
6. Conclusions
7. References
1. Introduction
Leptospirosis was first discovered by Adolf Weil as a disease 17th century (Bisen & Raghuvanshi, 2003), while reporting an “acute infectious disease with enlargement of spleen jaundice and nephritis.” Leptospirosis is caused by a spirochete Leptospira, which is primarily a zoonotic disease with an accidental human infection. Recent outbreaks of the disease indicated that it has become an emerging infection in human beings (Bisen & Raghuvanshi, 2003; T. Fraga, Barbosa, & Isaac, 2011). The disease usually appears wherever humans come in contact with the urine of infected animals as walking in a urine polluted environment (Schuller et al., 2015).
The disease is the widely distributed zoonosis worldwide it is present in all continents except Antarctica and evidence for the carriage of Leptospira has been found in virtually all mammalian species examined (Van de Werve et al., 2013). Humans most commonly become infected through occupational, recreational, or domestic contact with the urine of carrier animals, either directly or via contaminated water or soil (Bisen & Raghuvanshi, 2013).
Leptospira are thin, helical bacteria classified into at least 12 pathogenic and 4 saprophytic species, with more than 250 pathogenic serovars (Adler & Pen, 2010). These organisms are obligate aerobes with an optimum growth temperature of 28–30 degree centigrade. They grow in simple media enriched with vitamins B1 and B12, long-chain fatty acids, and ammonium salts (Penna et al., 2017). Growth of leptospires is often slow on primary isolation, and cultures have to be retained for about 13 weeks before being discarded. Agar may be added at low concentrations (0.1–0.2%). In such semisolid media, growth reaches a maximum density in a discrete zone beneath the surface of the medium, which becomes increasingly turbid as incubation proceeds. This growth is related to the optimum oxygen tension and is known as a Dinger’s ring or disk. Leptospiral cultures are maintained by repeated subculture or by storage in semisolid agar containing hemoglobin. Long-term storage in liquid nitrogen also yields good results and is the preferred method of storage for maintaining virulence(Girault, Soupé-gilbert, Geroult, Colot, & Goarant, 2017).
The disease is a systemic disease of humans and domestic animals, mainly dogs, cattle and swine, characterized by fever, renal and hepatic insufficiency, pulmonary manifestations and reproductive failure. Clinical signs are quite variable; most cases are probably in apparent and associated with host-adapted serovars such as Canicola in dogs, Bratislava in horses and pigs(Buyuktimkin & Saier, 2016).
2. Leptospira at Genomics Level
Leptospira serovars are usually classified as Leptospira interrogans, Leptospira borgpetersenii, Leptospira kirschneri, Leptospira noguchii, Leptospira meyeri, Leptospira weilii, or Leptospira santarosai etc. Analysis based on 16S ribosomal RNA (rRNA) gene analysis shows that these serovars are found in one of the three phylogenic clades into which the genus can be divided.
The genome comprises two circular chromosomes, CI (approximately 4.3 Mb) and CII (approximately 350 kb).They have a relatively large genome (>3.9 Mb) when compared to other spirochetes, (1.1 Mb) and Borrelia burgdorferi (1.5 Mb) (Buyuktimkin & Milton, 2015), characterized by a G+C content of 35–42 mol%, depending on the species, with a genome size ranging between 3.9 and 4.6 Mbp (Table 1). There are between one and two of each of the rRNA genes in Leptospira. In contrast to the situation in many other bacteria where the 16S (rrs), 23S (rrl), and 5S (rrf) rRNA genes are clustered and co-transcribed, those in Leptospira are widely scattered on the large chromosome Slow-growing pathogenic species L. interrogans and L. borgpetersenii and the faster growing intermediate L. licerasiae and saprophyte L. biflexa have a similar number (35–37) of transfer RNA (tRNA) genes (T. Fraga et al., 2011).
The genome of L. borgpetersenii serovar Hardjobovis strain L550 comprises two circular chromosomes of 3,614,446 bases and 317,336 bases, with an overall guanine and cytosine (G+C) content of 41.3%. The density of CDS sequences across the genome is 80.3%, with an average gene size of 931 bases. In total 3111 and 292 CDS features were annotated on chromosome 1 and chromosome 2, respectively. The L. borgpetersenii genome is smaller than the L. interrogans genomes and codes for proportionally fewer genes. The G+C content is higher than that found for L. interrogans, consistent with previous estimates of G+C content.
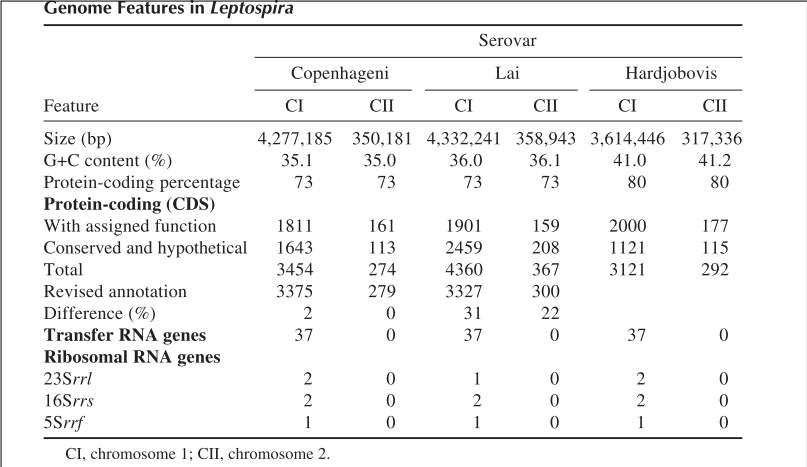
Table 1. Essential features of the genomes of pathogenic and saprophytic Leptospira spp. (Xue, Yan, & Picardeau, 2009).
Several insertion sequences such as IS1500 and IS1533 have been identified in Leptospira. The copy number of these IS elements varies considerably between serovars and between the isolates of a given serovar. The recent sequencing of Leptospira genomes allowed the identification of several other IS elements, one example being. These IS elements belong to a diverse range of IS families, including IS110, IS3, and IS4 The number of insertion sequences in L. borgpetersenii (mostly from the IS110 family) is much higher than those in L. biflexa, L. licerasiae, and L. interrogans Genome reduction in L. borgpetersenii serovar Hardjo may be the result of genomic deletions or rearrangements mediated by IS elements. A few phages related genomic islands have been characterized in L. interrogans and L. licerasiae, but many more laterally acquired regions can be detected in the genomes of Leptospira.
Although nothing is known about phages that may infect pathogenic Leptospira, it was shown previously that one of these genomic islands can excise from the L. interrogans chromosome and form a circular. In the saprophytes, the genome of the temperate leptophage LE1, with morphology similar to group A1 in the family Myoviridae was fully sequenced. The 74 kb LE1 prophage, which can replicate as a circular replicon in L. biflexa, has a GC content of 36 %, similar to that of Leptospira spp. Most of the 79 predicted coding sequences (CDS) display no similarity to known CDS, but 21 CDS appeared to be organized in clusters that might encode head and tail structural proteins and immunity repressor proteins(Picardeau, 2015).
The L. borgpetersenii genome is approximately 700 kb smaller and has a lower coding density than L. interrogans, suggestive of decay through a process of IS-mediated reduction of the functional genome. Additionally, many genes which are intact in L. interrogans have accumulated disruptive point mutations in L. borgpetersenii. Significantly, the loss of gene function is not random, but is centered on impairment of environmental sensing and metabolite transport and utilization. These features distinguish L. borgpetersenii from L. interrogans, a species with minimal genetic decay, and which survives extended passage in aquatic environments encountering a mammalian host. On the other hand, L. borgpetersenii survives poorly in the environment and it was concluded that it is in the process of evolution towards dependence on a strict host-to-host transmission cycle (Adler & Pen, 2010; Evangelista & Coburn, 2010; Mohammed, Nozha, Hakim, Abdelaziz, & Rekia, 2011).
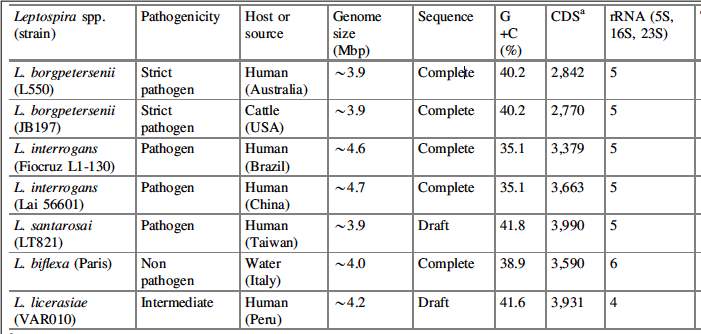
Comparative genomics of the 2 pathogenic and one saprophytic species has identified 2052 genes common to all, the core leptospiral genome (Fig. 1), a finding consistent with a common origin for leptospiral saprophytes and pathogens. Genome comparisons also allow the identification of pathogen-specific genes. The overall proportion of genes encoding proteins of unknown function is $40%. However, genes of unknown function are over represented in the genes unique to pathogenic species. Of 627 genes unique to L. interrogans, more than 500 (80%) encode hypothetical proteins. For L. borgpetersenii more than 200 out of 265 (75%) unique genes encode proteins of unknown function. These data are consistent with the notion that Leptospira possesses unique virulence factors which cannot be identified by similarity to those of other bacteria.
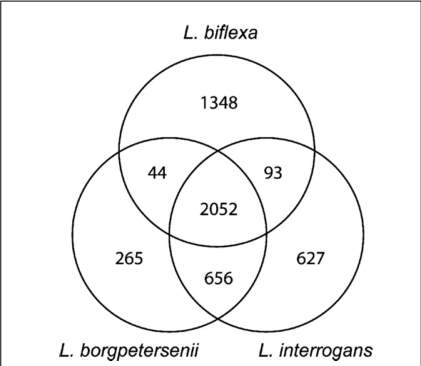
Fig. 1. Comparative genomics of Leptospira spp. Numbers of shared and unique genes amongst L. borgpetersenii, L. interrogans and L. biflexa. Adapted from(Adler & Pen, 2010).
3. Pathogenesis
Although the pathology associated with leptospirosis has been well described for decades, the pathogenesis of leptospiral disease remains to be elucidated. A diffuse vasculitis is thought to be responsible for many of the clinical manifestations. Disease onset is variable, taking place within one day to four weeks after exposure (Evangelista & Coburn, 2010).
3.1 Pathogen Entry and invasion
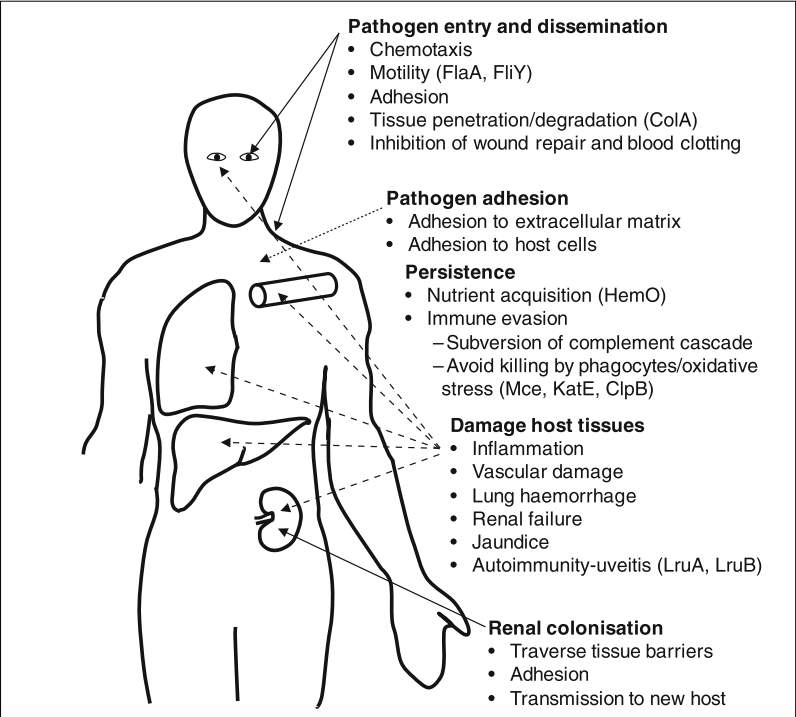
Leptospires enter the body through small cuts or abrasions, via mucous membranes such as the conjunctiva or through wet skin. They circulate in the blood stream, with the bacteremic phase lasting for up to 7 days. After the numbers of leptospires in the blood and tissues reach a critical level, lesions due to the action of undefined leptospiral toxin(s) or toxic cellular components and consequent symptoms appear (Adler & Pen, 2010).
3.2 Virulence factors
The virulence factors of Leptospira spp. discovered to date are either surface-exposed proteins that are primarily involved in attachment and invasion of host cells or are secretory proteins that either form pores or cause damage/lysis of host cells. Surface proteins of Leptospira spp. play important roles in attachment to host fibroblasts, microglia, endothelial and epithelial cells. Several surface proteins of Leptospira spp. have been identified that have been shown to bind to various components of extracellular matrix (ECM) and thereby mediate host-pathogen interaction. Some of the important virulence factors involved in leptospiral invasion and pathogenesis are discussed below.
3.2.1 Lipopolysaccharide (LPS)
LPS is considered to be a general virulence factor of Gram-negative bacteria. It is a major component of the outer membrane and contributes to its structural integrity. Leptospiral LPS has a composition similar to that of other Gram-negative bacteria but has lower endotoxic activity. LPS is responsible or antigenic diversity of pathogenic Leptospira spp. due to variation in the O antigen among different serovars, thus serving as the key molecule for diagnostic tests.
3.2.2 Loa 22
A lipoprotein with a peptidoglycan-binding motif similar to that of outer membrane protein A (OmpA), Loa 22 was the first genetically defined leptospiral virulence factor. Loa 22 is highly conserved among pathogenic leptospires, the mutant of which is avirulent in both the guinea pig and hamster models of leptospirosis. Complementation tests were found to restore virulence. Loa 22 is a surface-exposed protein that is recognized by sera from human leptospirosis patients. Although Loa 22 binds to laminin, the interaction is weak. The saprophytic L. biflexa species shares an ortholog of Loa22, but differential expression and variation in sialylation of this molecule in pathogenic leptospires could be responsible for its role as a virulence determinant
3.2.3 LipL32
Also known as hemolysis-associated protein I (Hap I), LipL32 accounts for 75% of the outer membrane proteome of leptospires. This lipoprotein is surface exposed and highly conserved in pathogenic Leptospira spp. LipL32 induces tubule interstitial nephritis through a TLR2-dependent pathway followed by activation of nuclear transcription factor kappa B, mitogen-activated protein kinases, and differential induction of chemokines and cytokines relevant to tubular inflammation.
3.2.4 LenA
LenA, also known as Lsa24, is a 24-kDa protein that binds to laminin as well as complement factor H, fibrinogen and fibronectin. LenA is member of the leptospiral endostatin like protein family that includes LenB, LenC, LenD, LenE, and LenF, all of which bind to fibronectin. Other laminin-binding leptospiral proteins include Lsa21, Lsa27, Lsa63, and a 36 kDa membrane protein
3.2.5 Hemolysins
The genome of pathogenic Leptospira spp. encodes various putative hemolysins and proteases that may facilitate the entry of Leptospira spp. into host cells and rapid translocation into various tissues. These include pore forming proteins and sphingomylinases. Hemolysins have the ability to lyse erythrocytes and other cell membranes, which make them potential virulence factors.
3.3 Adhesion and persistence of Leptospira spp. in the host
Leptospira spp. are not facultative intracellular pathogens like Salmonella or Mycobacterium but reside within host cells only transiently as they rapidly translocate through cell monolayers in vitro. Within non-phagocytic host cell, they reside either in the cytoplasm or phagosomal compartments.
There are several factors that contribute to leptospiral persistence and immune evasion in the host.
Following infection, pathogenic leptospires appear in the blood and invade almost all tissues and organs of the host(T. Fraga et al., 2011). The host’s immune system starts clearing leptospires from blood and other tissues, but the organisms may settle in the convoluted tubules of the kidney thereby may be shed in urine for a few weeks or even longer. Even after they are cleared from kidneys and other organs, they may persist in the eyes for a longer duration(Lehmann, Matthias, Vinetz, & Fouts, 2014).
3.4 Mechanisms of Damage to Host Tissues
The mechanisms by which leptospires cause host tissue damage and disease are not well defined. In particular, the molecular and cellular basis for virulence remains poorly understood, due mainly to the absence, until recently, of genetic tools for the manipulation of Leptospira. (Buyuktimkin & Saier, 2016; Martins & Silva, 2017).
They are characterized by various symptoms, including vasculitis, acute renal failure, jaundice, thrombocytopenia, pulmonary hemorrhage, myocarditis, conjunctival suffusion, and uveitis. The mechanisms by which damage occurs are not conclusively known. Injury to the endothelium of small blood vessels may contribute to ischemia and dysfunction of multiple organs, while circulating toxic cellular components or undefined toxins may contribute to tissue damage. Disruption of tissue integrity may occur by activity of leptospiral sphingomyelinases and phospholipase D. Leptospires also encode multiple proteases that may damage host tissues (col- lagenase, metalloproteases, and multiple thermolysins. Activation of host proteases such as plasminogen and matrix metalloproteases may also contribute to host tissue destruction and bacterial dissemination. Hemorrhage may be a result from a combination of tissue damage, disruption of hemostatic mechanisms, and interference of wound repair. Fever is a key feature of leptospirosis, and many pathologies associated with leptospirosis may result from inflammation. Inflammation may be a consequence of tissue damage rather than be directly mediated by bacterial factors.
Renal pathology during leptospirosis is associated with interstitial nephritis and cellular infiltrates containing neutrophils and monocytes, suggesting an inflammatory mechanism. Uveitis is another complication of leptospirosis that may occur weeks to years after initial infection. The condition may result from a breakdown in the immune privilege status of the eye with a combination of inflammation and autoimmunity
4. Antigen and Immune Response to Leptospira infection
Pathogenic Leptospira species are able to survive, and are more resistant to the action of the complement system, especially if they are virulent. The term pathogenic refers to genotypic properties of leptospires that may be expressed or not whereas the term virulent refers to phenotypic characteristics by which means pathological changes may be affected in the host. For example, L. interrogans serovar Copenhageni causes severe disease in humans when expressing its virulence mechanisms, which can be lost to a variable extent, in culture, thus becoming relatively a virulent, but not non-pathogenic. By contrast, L. biflexa serovar Patoc does not cause disease in any known host, thus receiving the classification of non-pathogenic. The property of virulence may be re-established by passage through a suitable host system (Lehmann et al., 2014).
4.1 Host Detection of Pathogen
4.1.1 Pattern Recognition Receptors
Mammalian cells display a variety of receptors on the cell surface with the design and purpose to recognize molecular signatures that are characteristic for microbial pathogens; these signatures are referred to as pathogen-associated molecular patterns (PAMPs). PAMPs include a wide variety of molecules such as bacterial LPS, lipoproteins, peptidoglycan, and flagella proteins. The host receptors, referred to as pattern recognition receptors (PRRs), interact with PAMPs, initiating a series of intracellular signals that trigger the host response to infection. Host receptors that recognize PAMPs include toll-like receptors (TLRs) and C-type lectin receptors (CLRs). A related group of receptors that recognize damage associated molecular patterns (DAMPs), which develop during infection, include receptors for advanced glycosylation end products (RAGE). Nucleotide- binding oligomerization domain (NOD)-like receptors (NLRs) recognize both PAMPs and DAMPs.
4.2 Innate Immunity against leptospiral infections
The innate immune system constitutes the first line of host defense, playing a crucial role in early recognition and elimination of leptospires. The activation of the alternative pathway of the complement system is one of the most important effector mechanisms during the first hours of infection (Polle et al., 2014).
Complement is a key component of the innate immune system responsible for protection against pathogenic microorganisms. Pathogens have evolved strategies to escape clearance by complement, either by sequestering host complement regulators or by down regulating complement activation. Virulent strains of Leptospira are resistant to human sera complement killing, while the saprophyte L. biflexa are sensitive (Fernandes et al., 2016).
4.2.1 Cellular Response
Monocytes/macrophages utilize PRR-mediated activation to provide innate immune protection of the host, especially during the early stages of infection. Leptospiral LPS and hemolysins stimulate macrophages to produce IL-1β, IL-6, IFN, and TNF- α in addition, macrophages treated with leptospiral LPS have enhanced phagocytic activity. The role of antibody appears to be important for macrophage-mediated killing of Leptospira.
Leptospires are readily phagocytosed by macrophages and neutrophils, both in vitro and in vivo in most animal species. In addition, antibodies readily lyse leptospires in the presence of complement. Polymorphonuclear leukocytes (PMNs) include granulocytes, eosinophils, and neutrophils, and these cells comprise important components of the innate immune response, but their role in protection against Leptospira infection is unclear. Two antibacterial peptides produced by bovine neutrophils, (Castiblancovalencia et al., 2016; Rodrigues, Santos, Lima, Gomes, & Brites, 2016).
4.3 Acquired Immunity
4.3.1 Humoral Mediated
Immunity in leptospirosis is predominantly humoral mediated in humans and most animal species, including dogs, pigs, guinea pigs and hamsters (Chen et al., 2013). Evidence for this arises from many findings showing that immunity can be passively transferred by convalescent human or animal serum, by experimentally produced antiserum, or by appropriate monoclonal antibodies (Mabs) such as those directed against leptospiral LPS which have been shown to protect guinea pigs, hamsters, dogs and monkeys against lethal (Lehmann et al., 2014).
As leptospires are extracellular pathogens, the acquired immune response depends on the production of antibodies and the activation of the classical pathway of the complement system. In leptospirosis, most of the specific antibodies produced are against the LPS. As a consequence, passive immunization with polyclonal or monoclonal anti-LPS antibodies is able to confer protection against the disease. In many experimental models, it became evident that phagocytosis of Leptospira by neutrophils and macrophages is only effective if this pathogen is opsonized by specific IgG. Besides opsonization, these antibodies may agglutinate leptospires and activate the classical path- way of the complement system. (Fernandes et al., 2017).
Antibodies against Leptospira have a key role in providing immune protection against lethal infection in many potential host species.
A key Leptospira antigen that is important for the development of immune protection in many host species is LPS. Immunization with Leptospira LPS protects hamsters against homologous challenge. Passive transfer of antibody to LPS has also been used successfully to protect mice, guinea pigs, monkeys, and dogs before lethal infectious.
Indeed, development of antibody to the LPS component of whole cell bacterins is thought to be key for immune protection against lethal infection with several Leptospira serovars in many animal species.
4.3.2 Cell-Mediated Immunity
Interestingly, although pathogenic Leptospira is not considered a typical intracellular pathogen, it was reported that L. interrogans may be able to escape from the phagolysossome to the cytosol of a human macro- phage cell line (THP-1). Therefore, leptospiral peptides could be complexed with MHC class I molecules, and be later presented to CD8+ T lymphocytes. Indeed, CD8+ T lymphocytes specific to peptides derived from leptospiral immunoglobulin-like (Lig) A protein were identified in human patients (Fernandes et al., 2017).
Both B and T lymphocytes have important roles in promoting an immune response to Leptospira infection. B cells appear to be primarily responsible for IFN-γ production and bacterial clearance in the liver, whereas T cells are responsible for these roles in the kidney. Furthermore, histological evidence of kidney tissue damage is greater in CD3−/− animals lacking functional T cells, as compared to WT or μMT (B cell deficient) mice.
γδ T cells are a unique class of CD4− CD8− T cells that comprise approximately 30 % of the normal adult ruminant PBMC population (Hein and Mackay 1991). This unique class of T cells comprises a smaller percentage of PBMCs in nonruminant species, and is not as well characterized as CD4+ and CD8+ αβ T cells. Most bovine γδ T cells possess the WC1 scavenger receptor on the cell surface.
5. Vaccines against Leptospira
Vaccines for humans and animals have been used since the 1920s; almost all of them were prepared from whole leptospiral cells killed by a variety of methods, including heat, formalin, phenol, irradiation etc. The use of undefined, live a virulent, attenuated, or saprophytic leptospires has not gained acceptance. Many of these early preparations were too reactogenic for widespread human use. Attempts to reduce reactogenicity have included the use of protein-free media for growth of leptospires and the use of sub-cellular fractions, the active component of which was almost certainly LPS. Nevertheless, human vaccines containing whole killed leptospires have been used successfully in China following floods resulting in exposure of large populations to risk of leptospirosis and in Japan and Vietnam(Adler & Pen, 2010).
Vaccination of livestock can reduce urinary shedding and risk to human handlers especially when accompanied by appropriate education programs, awareness, and hygiene in the community, and support from the authorities responsible for administration of human and veterinary public health. Commercial Leptospira vaccines are available globally for cattle, pigs and dogs but vaccination has proved to be only partially effective, due in part to the serovar restricted nature of vaccine induced immunity and the potential presence of local serovars others than those included in the vaccines. A successful vaccination program requires continued epidemiological studies to assess the incidence of different Leptospira serovars in a given population (Balakrishnan & Roy, 2014).
Most vaccines developed in the past have consisted of whole cell bacterins containing leptospires inactivated by chemical means or heat. Very early studies by Broom showed that hamsters could be protected with phenol treated Canicola cultures, while Brunner and Meyer immunized dogs and hamsters with a bacterin containing Canicola and Icterohaemorrhagiae and showed only serovar-specific protection. Most bovine and porcine vaccines contain at least serovars Hardjo and Pomona respectively; in North America, commercial vaccines also contain serovars Canicola, Grippotyphosa, and Icterohaemorrhagiae. Protection against Hardjo infection in cattle has been suboptimal, for this reason, cattle are vaccinated up to every three months in areas of high leptospirosis prevalence (Draayer, Bruckner, Peña-moctezuma, & Srinivas, 2013).
Canine vaccines generally, but not exclusively, contain serovars Canicola and Icterohaemorrhagiae (Wiggans, Wennogle, Curtis, Chandrashekar, & Lappin, 2014). Vaccines generally protect against disease and renal shedding under experimental conditions, but transmission of serovar Icterohaemorrhagiae from immunized dogs to humans has been reported. Moreover, immunized dogs may be infected with serovars other than those contained in commercial vaccines. Recent vaccines may also include serovars Grippotyphosa and Pomona in addition to the traditional vaccine strains, in response to the increasing incidence of canine infection with these serovars. Leptospiral vaccines for pigs usually include one or more of serovars Pomona, Grippoyphosa, Bratislava, Canicola and Icterohaemorrhagiae. Serovar Bratislava has been reported as abortigenic for sows. Other serovars,e.g.Hardjo, do not elicit significant titers in vaccinated sows (Adler & Pen, 2010).
Existing animal vaccines are generally polyvalent from serovars common to the species and particular to the locality of interest. This is a useful approach for animals but impractical for humans who are exposed to numerous serovars through occupation and travel. Moreover, there appears to be variance not only by region but over time making a broad spectrum vaccine desirable but challenging. Vaccination has apparently been successful following flooding in China, Vietnam, and Japan. Vaccination of the general public, however, where the incidence of infection even during epidemics may actually be low, may not be necessary, especially since yearly revaccination is recommended. Further, evaluation of new vaccines will be challenging due to low attack rates and variation in infecting serovars both geographically and temporally (Balakrishnan & Roy, 2014; Fernandes et al., 2017).
5.1 Vaccine design strategies for Leptospira
Vaccines can conceivably be used as a prevention measure through immunization of humans or the reservoirs that transmit leptospirosis to humans. Bacterin vaccines have been used for years in the veterinary field. These vaccines do not confer cross-protective immunity to serogroups that are not represented in the vaccine.
There is currently no widely used vaccine for leptospirosis. The first vaccine introduced for leptospirosis was a killed whole cell vaccine that consisted of formalin-killed leptospires. Various studies report the duration of efficacy of whole cell vaccines to be between 6 months to 7 years. However, in most studies, the duration of protection was at best 3 years. The problem with this vaccine is that its serovar specific. The monovalent vaccine did not protect against infection by other serovars and therefore its protection is dependent on the locally isolated serovars. This fact, plus its side effects, has led to other options being explored in vaccine designing. Leptospiral lipopolysacharides (LPS) are an area of interest for vaccine developers. However, immunity generated by these antigens was also considered to be serovar specific.
Overall, despite the advances in biotechnology, the only usable efficacious vaccine for leptospirosis to date are the whole cell inactivated vaccines. Vaccines based on recombinant membrane proteins have only been tried out in animals with limited success. The disadvantage of whole cell vaccines is that they are serovar specific (poly valent vaccines can be made by using several serovars in one vaccine) and therefore can be used in a geographically restricted area. Nonetheless, given the limited progression on developing a universally useful vaccine active against all pathogenic serovars, the most cost effective measure for a developing country is to work on a locally effective killed whole cell vaccine.
5.1.1 Bacterin vaccines
A variety of methods were used to kill leptospires for use as vaccines, including heat, formalin, phenol, ethanol, freeze-thawing, and irradiation. However, in the past 100 years very little has changed and at the present time the only licensed vaccines remain whole cell, inactivated bacterins. These have been used widely in cattle, swine, and dogs; specific usage for individual animal species is detailed in chapter by W.A. Ellis, this volume. Because of problems with reactogenicity due to components of the leptospires and constituents of the growth media, bacterin vaccines for humans have not gained acceptance to the same degree as for animals. Attempts to overcome these problems have included the development of protein-free growth media immunity elicited by bacterin vaccines is restricted to serovars with relatedagglutinating antigens and is generally humorally mediated, with the exception of Hardjo infection in cattle.
This restriction, therefore, requires a good knowledge of the regional epidemiology, which can be reliably gained only by culture and identification of locally prevalent serovars. Reliance on serological surveys to predict local serovars is not recommended. A second consequence of the limitation of immunity to serologically-related serovars is that in situations where multiple serovars are circulating, multivalent vaccines are required. Accordingly, bacterin vaccines containing up to four serovars are commonly used in many countries, especially in dogs and pigs (see the chapter by W.A. Ellis, this volume). Claims for protection against additional, closely-related serovars are probably valid. In some countries, locally-produced vaccines, especially for use in cattle, may contain up to eight serovars. Efficacy studies to demonstrate protection against all the included serovars have very rarely, if ever, been performed. Any possible antigenic competition effects between such large numbers of serovars are completely unknown.
A final drawback of bacterin vaccines arises from the fact that the immunity elicited is directed mainly against the leptospiral lipopolysaccharide (LPS), a T-independent antigen, and therefore involves IgM antibodies and lack of a memory response. The duration of immunity is therefore relatively short, with annual vaccination recommended in almost all cases.
5.1.2 Live Vaccines
The lack of detailed knowledge about leptospiral and the unavailability of genetic tools for easy manipulation of pathogenic Leptospira spp. have to date precluded any development of rationally attenuated, live vaccines. Nevertheless, serovar Pomona attenuated by laboratory passage was used as a live vaccine and shown to elicit protective immunity in hamsters, swine, and cattle. Although demonstrated to be safe and to be effective in stimulating a duration of immunity which lasted at least 14 months.
5.1.3 Lipopolysaccharide Vaccines
Unlike some of the other major spirochete genera Treponema and Borrelia, the major surface component of Leptospira is LPS. Furthermore, leptospiral LPS is a protective antigen. Monoclonal antibodies against LPS can protect against acute lethal infection in guinea pigs and hamsters and also protected dogs, based on recovery of leptospires from blood.
5.1.4 Recombinant Proteins as Vaccines
The development of recombinant DNA techniques and the availability of leptospiral whole genome sequences have resulted in a resurgence of activity to identify protective antigens and to develop defined subunit vaccines. However, much of the vaccination work reported in the literature suffers from the use of inadequate challenge doses, lack of reproducibility, and inappropriate statistical analysis. Accordingly, many of the claims about protection from infection, especially those claiming partial protection, cannot be substantiated.
6. Conclusions
In conclusion, leptospiara is a highly pathogenic and zoonotic disease that needs further scientific based researches at genomic level in order to have a good understanding of the disease pathogenicity, virulence factors and immune response. As stated in section 5 of this review, there is no a well-developed vaccine for prevention of the disease in animals and humans. This may be partly due to the lack of study of the genomics of the organism. Hence, advanced molecular studies of the disease are a a timely action which may lead to a novel findings and in designing effective vaccines with a limited side effects and low cost.
7. References
Adler, B., & Pen, A. (2010). Leptospira and leptospirosis. Veterinary Microbiology Journal, 140, 287–296. https://doi.org/10.1016/j.vetmic.2009.03.012
Balakrishnan, G., & Roy, P. (2014). Comparision of efficacy of two experimental bovine leptospira vaccines under laboratory and field. Veterinary Immunology and Immunopathology, 159(1–2), 11–15. https://doi.org/10.1016/j.vetimm.2014.03.002
Bisen, P., & Raghuvanshi, R. (2003). Leptospirosis Introduction. In Emerging Epidemics: Management and Control (pp. 176–219).
Bisen, P., & Raghuvanshi, R. (2013). Leptospirosis. In Emerging Epidemics (pp. 176–219). CHAP, John Wiley & Sons, Inc. https://doi.org/10.1002/9781118393277.ch7
Buyuktimkin, B., & Milton, S. (2015). Comparative genomic analyses of transport proteins encoded within the genomes of Leptospira species. Microbial Pathogenesis, 88, 52–64. https://doi.org/10.1016/j.micpath.2015.07.019
Buyuktimkin, B., & Saier, M. H. (2016). Comparative analyses of transport proteins encoded within the genomes of Leptospira species. Microbial Pathogenesis, 98, 118–131. https://doi.org/10.1016/j.micpath.2016.06.013
Castiblancovalencia, M., Fraga, T., Breda, L., Vasconcellos, S., Figueira, C., Picardeau, M., … Isaac, L. (2016). Acquisition of negative complement regulators by the saprophyte Leptospira biflexa expressing LigA or LigB confers enhanced survival in human serum. Immunology Letters, 173, 61–68. https://doi.org/10.1016/j.imlet.2016.03.005
Chen, H.-W., Zhang, Z., Halsey, E. S., Guevara, C., Canal, E., Hall, E., … Ching, W.-M. (2013). Detection of Leptospira-specific antibodies using a recombinant antigen-based enzyme-linked immunosorbent assay. The American Journal of Tropical Medicine and Hygiene, 89(6), 1088–1094. Journal Article, Research Support, U.S. Gov’t, Non-P.H.S. https://doi.org/10.4269/ajtmh.13-0041
Draayer, H. A., Bruckner, L., Peña-moctezuma, A. De, & Srinivas, G. (2013). International regulatory requirements for Leptospira vaccine potency testing . Roundtable : Current requirements and opportunity for harmonization. Biologicals, 41(5), 305–307. https://doi.org/10.1016/j.biologicals.2013.06.008
Evangelista, K. V, & Coburn, J. (2010). Leptospira as an emerging pathogen: a review of its biology, pathogenesis and host immune responses. Future Microbiology, 5(9), 1413–1425.
Fernandes, L. G., Siqueira, G. H., Teixeira, A. R. F., Silva, L. P., Figueredo, J. M., Cosate, M. R., … Nascimento, A. L. T. O. (2016). Veterinary Immunology and Immunopathology Leptospira spp .: Novel insights into host – pathogen interactions. Veterinary Immunology and Immunopathology, 176, 50–57. https://doi.org/10.1016/j.vetimm.2015.12.004
Fernandes, L. G. V, Teixeira, A. F., Filho, A. F. S., Souza, G. O., Vasconcellos, S. A., Heinemann, M. B., … Nascimento, A. L. T. O. (2017). Immune response and protective profile elicited by a multi-epitope chimeric protein derived from Leptospira interrogans. International Journal of Infectious Diseases, 57, 61–69. https://doi.org/10.1016/j.ijid.2017.01.032
Fraga, T., Barbosa, A., & Isaac, L. (2011). Leptospirosis: Aspects of Innate Immunity, Immunopathogenesis and Immune Evasion From the Complement System. Scandinavian Journal of Immunology, 73(5), 408–419. JOUR. https://doi.org/10.1111/j.1365-3083.2010.02505.x
Fraga, T. R., Barbosa, A. S., & Isaac, L. (2011). Leptospirosis: Aspects of Innate Immunity, Immunopathogenesis and Immune Evasion From the Complement System. Scandinavian Journal of Immunology, 73(5), 408–419. JOUR. https://doi.org/10.1111/j.1365-3083.2010.02505.x
Girault, D., Soupé-gilbert, M., Geroult, S., Colot, J., & Goarant, C. (2017). Isolation of Leptospira from blood culture bottles. Diagnostic Microbiology & Infectious Disease, 88(1), 17–19. https://doi.org/10.1016/j.diagmicrobio.2017.01.014
Hartskeerl, R. A., Collares-Pereira, M., & Ellis, W. A. (2011). Emergence, control and re-emerging leptospirosis: dynamics of infection in the changing world. Clinical Microbiology and Infection, 17(4), 494–501. JOUR. https://doi.org/10.1111/j.1469-0691.2011.03474.x
Lehmann, J. S., Matthias, M. A., Vinetz, J. M., & Fouts, D. E. (2014). Leptospiral Pathogenomics. Pathogens, 3, 280–308. https://doi.org/10.3390/pathogens3020280
Martins, E. A., & Silva, J. B. Da. (2017). Resistance of mice to Leptospira infection and correlation with chemokine response. Immunobiology. https://doi.org/10.1016/j.imbio.2017.05.017
Mohammed, H., Nozha, C., Hakim, K., Abdelaziz, F., & Rekia, B. (2011). LEPTOSPIRA : Morphology , Classification and Pathogenesis. Bacteriology & Parasitology, 2(6), 6–9. https://doi.org/10.4172/2155-9597.1000120
Penna, B., Marassi, C. D., Libonati, H., Narduche, L., Lilenbaum, W., & Bourhy, P. (2017). Diagnostic accuracy of an in-house ELISA using the intermediate species Leptospira fainei as antigen for diagnosis of acute leptospirosis in dogs. Comparative Immunology, Microbiology and Infectious Diseases, 50, 13–15. Evaluation Studies, Journal Article. https://doi.org/10.1016/j.cimid.2016.11.004
Picardeau, M. (2015). Leptospira and Leptospirosis, Current Topics in Microbiology. https://doi.org/10.1007/978-3-662-45059-8
Polle, F., Storey, E., Eades, S., Alt, D., Hornsby, R., Zuerner, R., & Carter, R. (2014). Role of Intraocular Leptospira Infections in the Pathogenesis of Equine Recurrent Uveitis in the Southern United States. Journal of Equine Veterinary Science, 34(11–12), 1300–1306. https://doi.org/10.1016/j.jevs.2014.09.010
Rodrigues, T. C. S., Santos, A. L. Q., Lima, A. M. C., Gomes, D. O., & Brites, V. L. C. (2016). Anti- Leptospira spp . antibodies in Crotalus durissus collilineatus kept in captivity and its zoonotic relevance. Acta Tropica, 158, 39–42. https://doi.org/10.1016/j.actatropica.2016.02.006
Schuller, S., Francey, T., Hartmann, K., Hugonnard, M., Kohn, B., Nally, J. E., & Sykes, J. (2015). European consensus statement on leptospirosis in dogs and cats. Journal of Small Animal Practice, 56(3), 159–179. JOUR. https://doi.org/10.1111/jsap.12328
van de Werve, C., Perignon, A., Jauréguiberry, S., Bricaire, F., Bourhy, P., & Caumes, E. (2013). Travel-Related Leptospirosis: A Series of 15 Imported Cases. Journal of Travel Medicine, 20(4), 228–231. JOUR. https://doi.org/10.1111/jtm.12035
Wiggans, K. T., Wennogle, S. A., Curtis, K., Chandrashekar, R., & Lappin, M. R. (2014). Vaccine-Associated Leptospira Antibodies in Client-Owned Dogs Leptospira Vaccine Responses in DogsMartin et al, 789–792. https://doi.org/10.1111/jvim.12337
Xue, F., Yan, J., & Picardeau, M. (2009). Evolution and pathogenesis of Leptospira spp .: lessons learned from the genomes. Microbes and Infection, 11(3), 328–333. https://doi.org/10.1016/j.micinf.2008.12.007
Cite This Work
To export a reference to this article please select a referencing stye below:
Related Services
View all


Related Content
All TagsContent relating to: "Pharmacology"
Pharmacology involves the study of drugs and how they affect the body. A pharmacologist contributes to drug development by researching and testing how the body reacts to medication, and whether the medication can have a positive impact on the body in terms of fighting illness and disease.
Related Articles

DMCA / Removal Request
If you are the original writer of this dissertation and no longer wish to have your work published on the UKDiss.com website then please: