Musical Protolanguage Amongst Early Humans
Info: 16369 words (65 pages) Dissertation
Published: 13th Dec 2019
Neanderthal Rock’n Roll: A case for musical protolanguage amongst early humans
Introduction
Almost all animate organisms communicate both conspecifically and heterospecifically through a range of signals, such as echolocation in bats or stereopsis in monkeys, with the intentions to interact, warn and, ultimately, propagate. However, amongst the enormous wealth of calls, gestures and displays, only humans are uniquely attributed to employing a language system determined by grammatical and syntactic recursiveness (Deacon 1997).
Despite attempts to teach language to our closest living ancestors, linguistic competence has ultimately failed to pass on into the animal kingdom (Terrace 1979), highlighting the profound complexity involved in producing even the simplest of utterances. Yet, because it comes so naturally, it’s easy to overlook the sophistication of this tool “separating brute and man” (Muller 1861: 23). Ideas in the human mind emerge as words selected from the lexicon, which when weaved into syntactic rules become ready for release as fully grammatical sentences via articulatory motions in the vocal tract. This carefully coordinated symphony of impulses- the faculty of language- is mankind’s greatest wonder, though the precise nature of its origin remains a mystery. The idea that language simply arose ex nihilo is dismissed by evolutionary linguists as being fallaciously misleading. Certainly, human beings today have language, the question is exactly what is the evolutionary origin of language, and more to the point, how did it occur?
Linguists, psychologists, philosophers, neuroscientists and anthropologists studying the evolution of language disagree strongly about what it is that evolved, where it came from, how it evolved, when and where largely because of the lack of direct evidence. Language does not fossilise, thereby shrouding its ancestries as a tremendously precarious topic amidst an empirical ethos, “no theme in linguistic science is more often and voluminously treated with less profitable result in proportion to the labour expanded” (Jespersen, 1922:412). In fact, evolutionary linguistics was deemed so disreputable that in 1866 the Société de linguistique de Paris banned all discussions about the origin of language altogether, a prohibition which remained influential in academia until recently in the twentieth century.
Despite this revitalisation, there is scarcely more agreement today; although, amidst the polarised proposals, there is a consensus in that language must have evolved gradually in a succession of ever more complex communication systems used by our relatives and ancestors. Convincingly aligned with the laws of evolution, researchers invoke the intermediary stage of protolanguage as the catch-all term helping us “…to bridge the otherwise threatening evolutionary gap between a wholly alingual state and the full possession of language as we know it” (Bickerton, 1995, pp. 51).
Generally, there are at least two ways to get from a one word stage to a composite language- divided between those evoking protolanguage as ‘compositional’ in character (Bickerton, 2000; Jackendoff, 2002), and those who see it as ‘holistic’ (Wray, 2000).
However, there is another interpretation that has long been hypothesised but largely ignored, one which represents a powerful marriage of comparative data, evolutionary insight and a biological perspective on language. Musical hypotheses cite the substantial overlap between ‘design features’ of music and language, and since most of the differences are semantic rather than structural, we may have handled a fundamentally musical/linguistic technique (Brown, 2000; Mithen 2005; and Fitch, in press). Early writers had conceived of a link between music and language by adopting a multicomponent view of human communication, as far back as Condillac (1714-1780) who first posited a “primeval song-language” (Thomas, 1995, p.72), though it was Darwin (1871) who linked language origins with, not only animal cries, but also an explicitly musical stage of human communication: “did our language commence with singing- is this the origin of our pleasure in music- do monkeys howl in harmony?” (Barrett, 1987, p. 568).
At present, advancements of the scientific arsenal have let investigations into biomusicology to solicit an experimentally comparative approach between human language and those animal communication systems traditionally termed ‘song’. While the relevant selective pressures that bestowed Homo with protolanguage are wildly varied, from communicating between mother and infant (Falk, 2009), to gossiping replacing grooming (Dunbar, 1996), I believe that no one selective force (e.g., sexual selection) is wholly accountable to elucidate all aspects of human musicality. However, a combination of neurological, physiological and archaeological data promises a deeper understanding of the biology and evolution of both behaviours.
Our last common ancestor (LCA) within the animal kingdom was the chimpanzees from whom we phylogenetically split from some 6 million years ago. Protolanguage and its related functions certainly emerged after this, though estimates point towards vastly different periods, as far back as 5 million years ago during the australopithecine era (Pinker, 1994) or as recently as Middle Palaeolithic 100,000 years ago (Novel and Davidson, 1996). Our enormous timespan is a double-edged sword because although our deep progress is implied through many tell-tale signs, an incomplete record may mislead and further mystify our understanding. Hence why I disagree with Bickerton’s view of language evolution as a “pencil-and-paper field” (2007:525) since it is the duty of linguists to invoke an interdisciplinary approach based on various empirical means to shed light on the emergence of language and music.
The aim of this research is thus three-fold. First, I will consider the two principal theories seeking to explain the inception of protolanguage. Whether compositional or holistic, theories of language evolution must be within the mental and physical capabilities of early hominids, as well as able to account for how key features of language could have arisen in the first place (i.e. semantics, syntax and phonology).
Then, I will argue why musical protolanguage jettisons the strengths of the holistic claim, though its dual-nature is doubly supported and twice as plausible. The intimate relationship between music and language is integral to human social life, closely linked in the brain and comparable to the communication in our primate cousins, all the while offering their own adaptive contributions towards our present genus.
Lastly, I review musical protolanguage in lieu of our closest related ancestor, Homo Neanderthalensis, whose behaviours, lifestyles and cultural achievements offer the invaluable insights into understanding late linguistic capabilities, both in the name of Neanderthals’ and our own.
2.1 Compositional protolanguage
The most recognizable and influential line of debate is championed by Bickerton (1990) who first argued for a Compositional account of pre-linguistic capabilities, and coined the term ‘Protolanguage’ itself, to explain the early constructions leading towards fully-fledged language. Bickerton’s position is driven by a Generative view of human language which regards grammaticality as a system of rules generating hierarchical balance behind a language’s syntax. It is possible to modify the scope of a word through such rules (e.g. marking plurality for dog-s), though this linguistic creativity presupposes a form of conceptual ability not present in the animal kingdom. By this logic, Bickerton’s solution was a protolanguage lacking “most or all of the formal structural properties that characterize language”, so to lessen the cognitive prerequisites to communicate in our early ancestors. Instead, protolanguage was a succession of proto-words (nouns and possibly verbs; Heine and Kuteva, 2007) uttered slowly and separately, with semantic content mapped onto an arbitrary phonological form. Each of these proto-words represented a single concept or real-world entity to form a 1-1 correspondence in meaning, hence known as compositional protolanguage (CPL), with only a loose pragmatic relationship between them. Thus, the lack of hierarchical structure or syntactic binding meant that proto-words would be concatenated paratactically like “beads-on-a-string” (p, 106) on a purely ad-hoc basis, denying any obligatory relationships with one another (Tallerman, 2011). Nevertheless, proto-words were combinable like they are in modern language to extend a complex line of thought, “Languages combine lawfully and protolanguages combine lawlessly” (p, 41). This lawlessness, though, would have been relatively tame being capped at telegraphic speech so to avoid ambiguity between agents and themes in multiple-word utterances until the advent of syntax. CPL first emerged to match signals referentially to concepts, so to aid “cooperation for hunting, gathering, defense, gossip or deception” (Jackendoff, 1999: 237). In essence, Compositional Protolanguage in all its variations (Bickerton, 1990; Hurford 2002; Tallerman 2007) maintains that words emerged first, followed by syntax and phonology exaptationally evolving on the basis of pre-existing semantic/symbolic representations due to a steep rise in signal processing capacities.
2.1.1 Excavating for compositionality
The fundamental challenge facing evolutionary linguists is that language does not fossilize. Only skeletal remains entertain any inferences, and even then they hardly amount to direct evidence of protolanguage. Instead, Bickerton (1990;1995) invokes the theory of recapitulation to draw clues from present communication systems that resemble successive stages in the linguistic evolution of our remote ancestors, deftly summed in the quip “ontogeny recapitulates phylogeny” (Haekel, 1874). Bickerton invites us to consider ‘living fossils’, “types of communication used by modern humans today that are close to, but do not share all the features of fully-modern language” (Kirby, 2009: 674) so to approximate relevant features. As such, protolanguage is arguably plausible if remnants of intermediate stages are functionally used today. Bickerton (1990: p.105) provides three kinds of linguistic behaviors that resemble a plausible evolutionary trajectory from protolanguage to language: pidgin communication, telegraphic stage of child language acquisition and the language of trained apes. However, these assumptions largely overlook what had to be gained for protolanguage to arrive in the first place; as we shall see, assuming a “modern language minus syntax” essentially given in the last LCA is not a safe assumption.
In the case of children’s early speech, a repertoire of mainly nouns (Macnamara, 1972) followed by simple verbs and modifiers (Bornstein, 2005) used by 2-year-olds supposedly reflects the referential structure of compositional protolanguage. Young children build a lexicon that is essentially syntactically-free, though arrive at a multipart meaning that is greater than if the words were in isolation by combining them (see figure 1), until a “catastrophic” acquisition of syntax occurs around 3 out of an essentially non-syntactic state. For Bickerton, “child grammars” embody the sudden evolution of ancient syntax, but this interpretation clashes with the consensus amongst child-language experts who recognise a fast but gradual acquisition (Gleason, 2005) of syntax in a piecemeal fashion, either verb by verb (Tomasello, 2000) or morpheme by morpheme (Brown, 1973). Even nativists who posit innately given knowledge as the basis for child language acquisition (Chomsky 1969) acknowledge that comprehension precedes production, so Bickerton’s analogy is inappropriate.
- “Mummy eat” (actor-action)
- “Bad kitty” (quality-subject)
- “Baby chair” (possession)
Figure 1.Composite meanings in children’s two-word stage. Taken from McGregor (2015: 233)
A better argument concerns the way a low-structured language could transition to a high-structured language, analogous with the Creolisation transition assumed by Muhlhausler (1986). Bickerton raises the case of pidgin communication, typically formed in slave plantations, where adults with opposing linguistic backgrounds are brought together and must negotiate a lingua franca. Initially consisting of short strings of words, pidgins are organised in an unfixed way and likely to vary between utterances thus lacking the complexity of modern languages, such as inflection and subordination. For example, the early ‘South Seas Jargon’ that emerged from contact between Pacific Islanders and Europeans during the 19th century (Figure 2):
- “Only he got using all the same pigeon”. (Gillbert Islands, 1860)
- “Me saba plenty” (Gillbert Islands, 1860)
- “Canoe too little, by and bye broke- All man go away, canoe gone, very good me stop”. (Lifu, Loyalty Islands, 1850)
Figure 2. Pidgin communication before creolisation. Taken from Clark 1979: 37)
Bickerton cites the rapid transition between pidgins towards fully-syntactic Creoles, like in the case of contemporary Nicaraguan Sign Language (Senghas, 2005), to illustrate the growth of CPL. However, creole learners already are language-ready humans, so this phenomenon only illustrates “catastrophic syntax” in a glossogenetic sense.
Bickerton’s strongest claim concerns the behaviour of home-reared apes who communicate with carers through signs and gestures. This type of evolutionary process is cultural, not biological, since alingual apes lack our language faculty and its related neural toolkits, therefore their outputs are bootstrapped by cognition-highlighting what it takes for language to develop between us and them. Fundamentally, great apes have demonstrated competence with non-iconic signs by drawing and gesturing lexigrams in short word-strings. The lowland gorilla Koko possessed tremendous mental powers to converse via gesture, play practical jokes and even coin new modifier-noun combinations to refer to objects for which she lacks signs, e.g. “white tiger” for a toy zebra, and “quiet chase” for hide and seek (Patterson, 1980). Researchers have argued that these apes can marshal a proto-grammar (Eisenman, 2013: 24) because of their linguistic categories and the cognition it implies (see Figure 3). Though not to dismiss the magnitude of achievement in linking signs to referents, this behaviour could not resemble erectus-like protolanguage like Bickerton proposes. Critically, neither vocal control nor ‘Mitteilungsbedurfnis’ (the need to talk to others) characterise the ‘protolanguage’ of highly trained apes, so we are left with an incomplete picture of unshared elements between our species and enculturated apes. This ‘protolanguage’ proves the mindfulness of Great Apes, not their disposition to speak.
- “Barefoot head” (to describe a bald man)
- “Giraffe bird” (To describe an ostrich)
- “Fine animal gorilla” (to describe herself)
Figure 3. Semantic relations in Koko’s corpus. Taken from Joseph (2013: 161)
Indeed, what all these linguistic behaviours share is a minimal structure, limited inflectional morphology and reduced stylistic or socio-linguistic variability (Dessalles 2006), which certainly distinguishes them from fully-fledged language. Nevertheless, Bickerton’s examples are unconvincing since they hinge upon the fallacy that the behaviour of modern humans, possessing all the genetic and neural mechanisms for language readiness, may resemble a template for the brain and behaviour of an extinct, possibly silent, hominid (I take issue with any ‘protolanguage’ assigned to Erectus since its rudimentary respiratory and vocal control could not meet the sonic demands to be reliably communicate; Wynn, 1998).
Rather, Bickerton’s ‘living fossils’ of human development (i.e. rapid acquisition of lexicon, phonology and syntax) do not recapitulate evolutionary changes from chimpanzees to moderns (Gould, 1977; Raff and Kaufman, 1983), but actually represent recent selective pressures for early learning after adult language already existed. If, for argument’s sake, we were to accept a word-based protolanguage, albeit around Homo Antecessor at least ~1.2MYA, then it must justify the emergence of late grammatical categories below.
2.1.2 Transition towards compositionality
For protolanguage to grow beyond a simple set of unitary utterances, theories must attest the increase in structural complexity typical of modern language (Jackendoff, 2002; Hurford, 2003; Tomasello, 2003). What is typically referred to as the synthetic transition lends credence to the mechanism pervading cross-linguistic diachronic change today. In grammaticalisation, open-class nouns and verbs become ‘semantically bleached’ allowing for their rebirth as function words from content words (Trask, 1996; Sweetser, 1988), now explicitly marking intra-syntactic relationships. For instance, the transformation of the verb meaning ‘go’ commonly becomes grammaticalised into future tense markers, concurrently seen across unrelated languages like Basque, Quecha, Tamil and Zulu (Heine & Kuteva, 2002). To see how this is possible, below is an example in English from constructions with non-finite complements:
- I am going [to see Bill]. (Purposefully directed motion)
- I am [going to] see Bill. (Event in future, not motion)
- I am [gonna] see Bill. (Phonetic reduction and simplification)
Figure 4. Grammaticalisation in English. Taken from Smith (100)
It follows that such re-analyses, driven by inferential semantic reconstruction, is the unidirectional candidate motivating the transition from simple nouns to syntactic elements (Tallerman, 2011). A great deal of work has investigated the rise and ordering of grammatical constructions, so a scenario of syntactic evolution is likely to follow Heine and Kuteva’s progression (Figure 5).
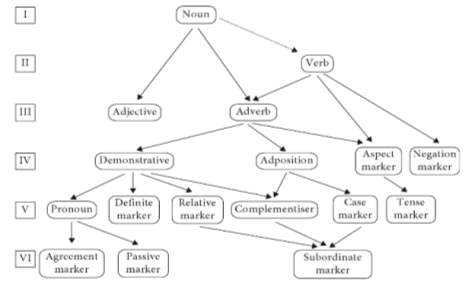
Figure 5. Layers of grammatical evolution (Heine and Kuteva 2007).
Whether syntax was integrated biologically, (Bickerton, 2009), or culturally (Tomasello, 1999; Jackendoff, 2002; Steels, 1997), it began the proclivity to arrange signals into larger, meaningful, syntactically recursive utterances, each with a specific meaning: “Once nouns and verbs came into being, well-understood linguistic processes will do the rest” (Tallerman, 2007: p18). The dichotomy of a cultural or biological constraint on syntactic structures is not strictly relevant to this paper, but one tested by interesting points (see Christiansen and Kirby, 2003 for a review). What is relevant, though, is to understand the precise means in which such constraints might play a role in language evolution. We must first understand the signalling mechanism used to produce linguistic utterances, an explanation overlooked by CPL which skips over the production/acknowledgment of linguistic signs in a prehistoric context, to instead concentrate on the emergence of composite phrases.
So, while the ‘words without grammar’ hypothesis compellingly draws upon linguistically-mediated behaviours, it does not overall satisfy the component transformation into FLB (Chomskyan term referring to our systems of sound, sign and meaning) because it presupposes an ability for vocal imitation and drive for referential communication; the existence of which cannot simply be accounted for ipso facto– all mechanisms preceding language require selective propulsion in functional succession. Hypotheses about hominine evolution based on paleontological evidence do not support hypotheses of an abrupt jump from ape-like vocalisations to the kind of protolanguage proposed by Bickerton (1990), and arguments about catastrophic syntax hinge upon debatable analogies, “Given that it is well known that apes in language training experiments can spontaneously adopt ordering, and even parrots can be trained to pay attention to the sequencing of symbols, it would be very surprising if our hominid ancestors did not share that skill” (Tallerman, 2007:21). It seems the inception of CPL is grounded upon a midway projection of progress, focussing only on “the end game of language evolution” (Fitch, 2010: 431), oversimplifying the acquisition of protowords by glossing over signalling stipulations.
The last decade has witnessed several amendments (see Hallam & Thaut, 2016, for a review), though this paper must move on to consider other works.
2.2 Holistic protolanguage
The pivotal dichotomy underlying investigations into our linguistic past splits into a compositional versus a holistic view of protolanguage (Bickerton, 1990; Wray, 1998; Tallerman, 2007; Arbib, 2005). Unsurprisingly, the avalanche of publications and discussions fuelling this debate have mystified the answer and even led to an entire issue of the Interaction Studies journal (Vol. 9:1, 2008) devoted to the idea of “Compositionality vs Holophrasis”, all firmly committed to unveil the way we first ‘spoke’.
Alison Wray (1998, 2002) and Michael Arbib (2002, 2005, 2008) are notable for proposing a holistic stage of language evolution whereby communication was realised as individual utterances expressing entire propositions. In effect, holistic protolanguage (HPL) was characterised by a “small inventory” of individual signals, lacking in internal morphological structure, which wholly convey “a complete communicative act” rather than being composed of semantic atoms (Arbib 2008: 154). Critically, our language was holistic because it developed from an earlier primate-like system of communication, characterised by vocal and gestural signals, designed for the “preservation of self, others and the servicing of and adjustment of relationships” (Wray, 1998, p.51). To clarify, human language did not directly evolve from the primitive communication of non-human primates, much like how humans did not directly evolve from apes, but rather HPL invokes primate communication for sharing properties like being holistic in nature devoid of discrete words or grammatical rules (Tomasello, 2003), akin to primate calls observable today (Mitani, 1996). It is helpful to imagine our ancestors yodelling alarm-signals similarly to how non-hominids alert conspecifics. For instance, when Capuchin monkeys feel threatened they emit high-pitched alarm calls (Fichtel et al, 2005) like their eagle alarm meaning “Beware of the eagle!”, or “Look up at the sky and run for cover!”. No part of the call means ‘beware’ or ‘run’, rather the entire call is the message. Following this logic, hominids initially communicated holistically via manipulatively-driven propositions to generate some form of desired behaviour in others, like commands, threats, greetings and requests. Only later in time did referential meaning via compositionality emerge when they were decomposed- or “segmented”- into more familiar word-like units (Wray, 2000).
For the ease of exposition, I will principally discuss Wray’s HPL (1998;2000) since subsequent conceptions of Holophrasis in protolanguage are underpinned by virtually identical principles. As we shall see, the intent to communicate selectively propelled hominids towards language, though the precise medium, whether gestural (Arbib, Corballis 2011, Pollick & de Vaal 2007) or musical (Mithen, Brown 2011, Fitch, 2009), each imposes their own specific constraints on the architecture of language and the brain.
2.2.1 Excavating for holism
Advocates of HPL reject the intermediate multi-word syntax-free stage of CPL. Instead, our ancestors used signals to represent entire propositions, each with an arbitrary form and complex meaning agreed on by the community. But how could human languages have evolved from primitive types of holistic communication? Like Bickerton (1990), Wray (2000) turns to ‘living fossils’, qualitatively different kinds of language lacking in modern syntactic complexity, as indirect evidence of language evolution to explain such continuity. While the synthetic model is motivated by communication in pidgins, young children and trained apes via grammaticalisation, the holistic model looks towards formulaic language and derives structure through segmentation.
Strictly speaking, the closest types of holistic utterances we have today are neologisms like ‘abracadabra’, which represents something like ‘I hereby invoke magic’ (Mithen, 2005). Nonetheless, by tapping into the conventionalised strings of exchanges, we uncover Holistic formulae which ubiquitously convey information as whole, inalienable chunks. Some formulaic utterances are manipulative since the speaker aims to invoke a physical or emotional reaction in the hearer: ‘watch where you’re going’; ‘I’m sorry’; ‘look out’; some are idiomatic: ‘Kick the bucket’ (whose meaning, ‘to die’, is arbitrarily related to form); or just regularly common constructions, ‘Was there anything else?’ (Wray, 2009). Indeed, these constructions can be analysed as containing individual words (only possible once a fully modern language with separate words already exists), but psycholinguistic models indicate that utterances with an agreed meaning are stored, retrieved and processed as if it were a single irreducible item without any necessity to activate grammar rules (Wray & Perkins 2000; Meyer 2015). For instance, we don’t consider parts ‘by’, ‘and’ and ‘large’ as words, but components of the adjunct ‘By and large’ which we unitarily manipulate. For Wray, the frequency of these formulae indicates a recurring disposition from an intermediate stage of holistic communication. The argument is that having no grammar implies no component words, annulling the possibility of novelty being spontaneously understood and shared by others. Thus, if we are reasoning about a language without grammar (like CPL), we should not start with words, but with holistic utterances with an agreed meaning.
Consequentially, the holistic processing strategy may predate the emergence of single referential words or the grammar that combines them. In time, segmentation was the process decomposing holistic utterances into component parts plus words which govern their combination (Wray, 1998; 297). In figure 6, Wray suggests example holistic utterances to illustrate how syntax may have emerged through chance co-occurrences of meaning and form.
- /mἐbita/ <—> (Give her the food)
- /ikatubἐ/ <—> (Give me the food)
- /madu/ <—> (Keep away)
- /mabu/ <—>(Take the stick)
- /tebimἐ/ <—> (Give that to her)
- /Kumἐpi/ <—> (Share this with her)
Figure 6. Characteristic holistic protolanguage utterances. Adapted from Wray (1998: 51).
Firstly, note how signals A & B share similar meanings, but do not bear any phonological resemblance to one another. Since they lack any recombinable units of meaning, their precise syllabic make-up and length is irrelevant: “within the bounds of memorisability and minimal differentiation, strings of any length and phonological structure may be used” (Wray, 2000: 294). The converse effect is demonstrated in signals C & D.
Secondly, such a set of phonologically distinguishable complex-signals supposes a markedly productive phoneme inventory. Early hominids would have modified their vocalisations into distinctive, recurrent syllable-sized chunks, which would have led to the gradual emergence of different vowels and consonants to set in place what MacNeilage (2007) calls “syllabic variegation”. Like the examples above, a wide range of distinct sounds (e.g. bilabial nasal /m/ to back voiceless velar plosive /k/, and maximally distinct cardinal vowels like high front /i/ to front low /a/) has the apparent advantage of explaining the anatomical evolution of early hominids’ vocal tract, whereby the generative potential of phonology now far outstrips modern linguistic needs.
Being an inherently functional system, speakers initially required only a small fixed set of messages to interact with fellow nomads. Out of this, presumably via ecological pressures, users developed the capacity to distinguish syllables by the segments governing their combination via an analytical route of segmentation. Chance co-occurrences of meaning and form led speakers to segment out words, discarding the rest of the residual template to yield nuggets of segmented meaning. Returning to Fig 6, consider how a learner exploits the partial regularity in messages E & F to conclude that the element /mἐ/ had the meaning: female person + beneficiary. The individual who discovers said regularity may fuse the meaning-to-morpheme correspondence and begin to produce novel utterances with it. Sometimes the hypothesis would be upturned in lieu of counterexamples, but sometimes the counterexamples would be ‘hypercorrected’ to incorporate /mἐ/, for example message C might alter to ‘Keep her away’. This process is consistent with Hockett’s (1960: p.38) speculation of why “duality of patterning” is a design feature of human languages (meaningful units within the system are created through combinations and re-combinations of sound units that have no meaning in themselves but which function to keep meaningful units distinct from one another).
The pressure to analyse utterances into their components and assign meaning likely came from an increasing memory load driven by an increasing repertoire of holistic utterances. With more signals becoming segmented, more must have phonologically complexified in response to communicative pressures for more lexical distinctions (Wells, 1987). What might have begun with contrastive monosyllabic utterances, our hominid ancestors improved by realising the creative potential of stretching words or reduplicating sequences rather than expanding the phonetic inventory, as in languages like Hawaiian with a small segmental inventory (10 atonal vowels and 8 consonants) (Mufwene, 2008) until a lexicon of words would result from fully analysed utterances with ordered relations. Development was gradual, possibly over the period that Homo Erectus came to vocalise (~1.8MYA) and Homo Heidelbergensis developed FLB (~700KYA), (McCarthy, 2015).
2.2.2 Discussion of Holism
Clearly, there is an enormous gulf between our language and that of our primate relatives. One approach is to determine what linguistic components are uniquely attributed to humans, i.e. recursive capacities (Hauser, Chomsky & Fitch, 2002; Pinker & Jackendoff, 2005); these are interesting but well beyond the scope of this chapter since we are primarily concerned with providing a backdrop of plausible protolinguistic systems compatible with musicality. More importantly, one must consider the origins of first meanings, which Wray (1998) appears to sidestep by invoking continuity with primate communication, though is indeed justified by drawing attention to the emotional exchanges of pre-linguistic primates. In the “smooth, gradual and beneficial” development towards protolanguage, the manipulative noises and gestures of our ancestors would need to become phonologically organised. Given enough segmentation time, Homo heidelbergensis and other early humans 700KYA could have had many holistic messages, ranging from ‘come-here’ to ‘go-and-hunt-some-fish-in-that-lake-we-bathe-in’, uttered and mutually understood to facilitate early co-operative lifestyles in the groups of ~150 hominids (Dunbar, 1993; Zhou, 2005). Unsurprisingly, HPL has received flak from critiques probing how credible this would be.
Firstly, the inception of HPL is criticised as too demanding a learning process for Homo erectus (the earliest species linked to protolanguage by Wray, Tallerman and Bickerton), “Assuming the input was a set of holistic utterances, could this feat conceivably have been matched, even approached, by the smaller-brained erectus…? I submit not” (Tallerman, 2007: 16). We could assume that the capacity for Homo erectus to memorise signals is the same as that of capable moderns (e.g. air-traffic controllers possess an enormous repertoire of coded signals), but Tallerman discounts learnability based on their relatively smaller brain size- some 300cc difference in cranial capacity. Yet for this to hold, there must be a relationship between brain size and capacity for lexical memorisation, a tentative link at best Jackendoff (2002). Our prelinguistic ancestors did not have the same mental capacity as modern Homo Sapiens, but the evolution that innovated phonetic communication did not then necessarily call for the kind of complex communication we engage in today.
Next, the transition from HPL to language via segmentation is arguably implausible because of surface instability across word-forms. Certainly, holistic expressions could not remain synchronically identical because phonological processes are ubiquitous: allophonic & allomorphic variations will diachronically emerge as sound change, “variation cannot help but exist because once hominids have a vocal tract in anything approaching its modern form, then specific phonetic tendencies appear spontaneously” (Tallerman, 2007:9). It would be unwise to deny the course of sound change, but segmentation does not require expressions to remain identical for learning. Chance similarities in an unfixed system are perfectly possible, just as it’s possible to win the lottery even if you change numbers with every draw. It would certainly take a long time to strike, but time is not an issue when dealing with millennia of evolution. In fact, a late acquisition of syntax is further justified by the exponential archaeological record, as we shall see (Chapter 4).
A better argument concerns the damage done by sound change: “How could the segmentation have proceeded successfully over hundreds of thousands of years when the material the speakers were working on was continuously slipping out of their grasp, changing the validity of any hypothesis formed by one generation and demolishing the emerging system?” (Tallerman, 2007:11). Could analysis based on regularity operate if sound change undermines the patterning? I believe the answer is evident across modern languages, all of which are messy, irregular yet orderly. In attested language change, analogical levelling reduces the number of allomorphs a form has so to spread uniformity across paradigms. Crucially, forms that have been damaged by sound change can be repaired into alternate regularities – seen in English ‘strong verbs’ with stem alternations (sing/sang/sung) which level to ‘weak verbs’ with an affix (bake/baked/baked) (Campbell, 1998: 106), all of which remain comprehensible. To see how analysis could proceed, computer simulations demonstrate how agents converge on co-ordinated communicative codes comparable to the evolution of natural language features. For instance, Brighton and Kirby (2001) show how holistic utterances may be learnt iteratively and become segmented despite destructive sound change.
Nevertheless, critiques of HPL mainly attack the continuity between the social, neural and vocal tract of “primate calls” and their basis in modern language. Tallerman’s (2011) objections are just (e.g. They are not intentional, they are produced with both outbreath and inbreath etc.) though these miss the virtue of HPL: it provides one step in an evolving system which overcomes precisely those differences. Moreover, I take issue with Tallerman’s repudiating views because if every capacity that has been exapted for speech had to remain the same, there would have been no evolution whatsoever. These objections overlook a principle tenet of evolution: something had to be co-opted from elsewhere, which in this case was the intentional production of vocalisations and their gradual modification into digital sequences (Wray, 2007). For the academic community to ever surpass scepticism, it must incite a Newtonian order of competing hypotheses and proceed with the most merited assumption, not refute ‘radicalism’.
At this point it would be appropriate to question the logistics of segmentation: what boundary conditions were suitable and if modern humans, with all their superior brain capacities, can segment at all? While evidence from historical linguistics (i.e. structure manifests when not previously present via back-formation and reanalysis; Simpson & Weiner, 1989) and child-directed speech (infants with limited attention and memory can segment in small templates; Lieven & Tomasello, 2003) suggests that humans only require few examples to segment, better direct evidence comes from Gomez (2002) who experimentally demonstrates how templates applying to highly variable fillers are more readily learnt by adults and children than a template which appears in more stereotyped circumstances.
Granted, the abilities of modern humans are not synonymous with the competencies of cognitively-inferior Homo to standardise co-occurrences of meaning and form, “words will never appear out of formulae unless the hominids using holistic protolanguage have both the necessary motor control and neural capacity to reorganise phonetic strings- how could these abilities exist prior to the language faculty itself?” (Tallerman, 2007: 595). To answer this, Saffran, Aslan & Newport (1996) demonstrate that in language acquisition, the segmentation of words from fluent speech can be accomplished by 8-month-olds based solely on the statistical relationships between neighbouring speech sounds, with as little as 2 minutes of exposure. Crucially, this mechanism that detects co-occurring segments across utterances is also present amongst Cotton-top Tamarins (the last Old World ancestor with Homo 40-million-years ago; Rosenberger, 1992) shown by Hauser, Newport and Aslin (2001).
It seems there are sufficient grounds to pair at least Homo Antecessor with the neural capacities for recognising phonetic strings, albeit in a rudimentary manner which might struggle with the complex hypothetical phonetic sequences proposed by Wray in Figure 6. To be certain, I believe that the initial phonetic distinctions, or ‘variegations’, were actually far simpler vocalic deviations, such as modifying the quality of a vowel or inserting a glottal stop, which could have been the tipping point to “…innovate [an] embryonic digital phonation system from continuous ape-like vocalisations” (Mufwene, 2008). A fragile developmental curve of trial and error, intensifying vocal control and increasing sociality could mark the evolutionary trajectory towards language today.
Interim CPL HPL
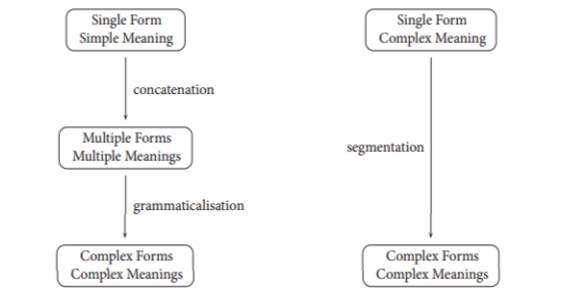
The path towards language was no doubt short of a miracle. Its very existence teases our every intuition, defies our conception of time and leaves us utterly enthralled by nature- meanwhile supposing the greatest inductive puzzle known to linguists. The culmination of countless works has led to a fork between the opening conditions for meaningful proto-forms, subsequently defining the character of protolanguage and the putative paths into modern language (see Figure 7).
Figure 7. Competing routes for the transformation of protolanguage into full language. (Taken from Smith: 101)
Since neither model is groundless and we cannot ask our ancient ancestors, it is the doctrine of evolutionary linguists to mould conjectures around powerful insights and fresh evidence, branching off into plausible variations upon the Holophrasis vs Compositionality debate.
One of these, the Musical Protolanguage approach (MPL), is an increasingly persuasive proposal linking music and communication under a Holistic outline. At its core, it judiciously tackles a major criticism at the forefront of the Holistic claim (Tallerman, 2007)- how fine, intentional articulatory control developed to produce a holistic signalling system; with MPL in mind, this criticism not only vanishes but ultimately strengthens the holistic camp.
Like language, music is an undeniable universal that reaches deep into our species’ past (Nettl, 2000) and continues to be significant across all human cultures (Cross, 2006). Again, music resembles language as an interactive and participatory medium (Small, 1998) that appears to constitute a communicative system (Miell, 2005), but one that is understood to only communicate emotions (Juslin and Sloboda, 2001). It seems strange then that a cognitively-expensive medium specialised in communicating emotion developed when language and gesture already powerfully accomplish this role (Arbib, 2005). The leading question, and indeed purpose of this paper, is to consider an evolutionary trajectory for language as having co-evolved from ancient musical vocalisations (Hurford 1998; Carstairs-McCarthy 1999; Knight 2000; Mithen 2009).
In essence, we speak together because we once sung together. Models of Musical Protolanguage must justify how core features of language, syntax and semantics, emerged from a ‘phonology-first’ approach to language evolution.
3 Music, Language and Evolution.
The inherent value of language is equivocal to the momentous achievements across mankind (tools, economy, society etc.) and remains an increasingly adaptive and functional tool today. Curiously, musicis an equally ancient and ubiquitous tool of human behaviour, presently stretching from the Eurovision Song Contest to the Amazonian Tsimané tribal-ceremonies, though its functional or adaptive motivations are unclear. Musical activities lack any obvious survival value in the individual, so why is music so pervasive amongst mankind? The relationship between these two behaviours is a contentious issue which may logically entertain several scenarios of evolution, but first we must carefully consider their similarities and differences to fully appreciate the subsequent adaptionist propositions of Musical Protolanguage.
3.1 Similarities and differences.
Cross-cultural and developmental research indicates that music and language have had a profound appeal to all modern and prehistoric species of Homo sapiens (Blackling, 1973). All cultures, in one way or another, make use of internal repetition and variation in musical and linguistic utterances to alter rhythmic structures based on distinctive lengths and dynamic stresses (Nettl, 1983). However, the public eye still discredits the universality of music by placing a greater emphasis on the production rather than reception, when in fact we are all equally musical in recognising variations in pitch, rhythm, tone and timbre, “society claims that only a limited number of people are musical, and yet it behaves as if all possessed the brain capacity without which no musical tradition can exist- the capacity to listen and distinguish patterns of sounds” (Blackling 1973: 47). After all, the flair in Beethoven’s symphonies only exists because of the audience’s ability to appreciate it.
On a structural level, music and language share the same nodes of expression: they can be manifest vocally, as in speech and song; physically, as in sign-language and dance; and in writing, as in the alphabet and musical notation. Moreover, the cultural transmission of a specific language or musical custom occurs by passive childhood acquisition, the production of which emerges through exchanges with experienced language/musical users. Both have a hierarchical structure, being constituted by acoustic elements (words or tones) that are combined into phrases (utterances or melodies), ultimately arranged by cultural-constraints (e.g. template morphology in the Arabic language or ternary form in Baroque compositions). Again, both forms operate on recursiveness- the embedding of a linguistic or musical phrase that enables a potentially infinite range of expressions to be generated from a suite of finite elements.
The fundamental differences emerge when considering how each system conveys their parts: in language, words are composed of symbols, but musical notes are not. Symbols allow words to carry an arbitrary relationship with the entity to which they refer (i.e. there is no direct connection between the shape and the concept); so, speakers of English have agreed and learnt that ‘bigot’ invokes the notion of an intolerant individual, but its arbitrariness also holds a cognate for ‘moustache’ in the Spanish lexicon. On the other hand, musical notes lack referential meaning because Middle C (261.63hz) only means Middle C. It could be argued that sequences of notes carry emotional meaning (e.g. the national anthem), but generally there is no agreed convention describing how notes bear semantic tokens within a “music community” as there is for words and meanings (Cross, 2001).
Linguistic utterances are generally compositional and governed by 3 rule systems: syntax, grammar of words; morphology, word-form combinations; phonology, relationship of speech sounds. These rules are useful in providing an extended meaning than what is just contained in the symbols themselves; hence, ‘Judy kicked Punch’ denotes something radically different than ‘Punch kicked Judy’, but reversing the order of notes would be less significant. Nevertheless, the abstraction of grammatical rules from the utterances heard comes remarkably easy for developing children, which could only be explained through an adapted set of predispositions for the acquisition of grammar among Homo Sapiens – ‘Universal Grammar’ (Chomsky, 1965). This innate awareness allows native speakers to identify forms that sound ‘wrong’, (e.g. dangling participle: ‘Where are you from?’ vs ‘From where are you?’), which might also apply to musical intuitions (e.g. modulations between cadences: II->V sound ‘Interrupted’ vs V->I sound ‘Perfect’). But attempts to justify the latter as an innate grammar of music (Lerdahl & Jackendoff, 1983) are unconvincing because such logic would confuse ‘innate grammars’ and systematicity of behaviours, like that in chess and computer tasks etc.
Only language is driven by intentional means to communicate referentially about any entity in the universe, “from soap operas to theories of the origin of the Universe, from promises to threats, from lectures to questions” (Pinker, 2003: 16); conversely, music lacks any clear communicative function (Cross, in press), but manipulates the profound emotional and physical states of interlocutors through entertainment (e.g. some might leave the opera wanting to dance, others might offer evaluation “I liked it”, and others might turn to metaphorical characterisation “it had the feeling of a long heroic struggle triumphantly resolved”; Sloboda, 1985:12).
Our existence is a product of evolutionary processes, a force that shaped our bodies and built our minds. Two of our wildest accolades are the abilities to bind sounds to referential meaning and manipulate musical modes, but how connected are these two behaviours? The discussion above is enough to treat them as separate, though their commonalities invoke a ‘chicken-and-egg’ dilemma in evolutionary provenance. Some have proposed that music is just a contingent by-product of evolution, something enjoyable but unimportant, whereas others view music as a major contributor in human evolution. More to the point, has musicality emerged as an adaptive feature for Homo?
3.2 Musicality: Adapted or non-adapted?
Most of the evolutionary accounts of musicology (Cross, 2009; Brown 2000; Bispham, 2006; amongst others) propose that musicality is a biological adaption of ancient evolutionary provenance, and thus we should show “developmental readiness for musical stimuli” (Fitch, 2010: 506): the clearest traces of adaption are evident in the capacities of the infant mind. A wealth of evidence shows that young children are sensitive to variations in rhythm, pitch, durational patterning and timbre (Trehub, 2003: 670), but most importantly, these competencies develop too quickly to be explained as the outcome of learning by interaction with the environment. In the same way that infants are primed for intuitive physics, expressed in their surprise at spatial violations (e.g. experimental magic tricks; Huntley-Fenner, 2002), the auditory system of newborns is tuned to discriminate between speech and non-speech stimuli (Voulomanous & Werker, 2007), because of dedicated cortical structures (Pena, 2003). Adults, and especially mothers, instinctively alter their prosody when talking to preverbal infants in ‘motherese’- a language characterised by higher/broader pitch and amplitude variation to capture infants’ attention and lead a causal role in language acquisition (Csibra, 2010:148). It is very likely that the universality of motherese amongst parents (Fernald, 1992), coupled with the infants’ sensitivity to its musical contours, are innate and adaptive since two-day-old newborns show a preference (Cooper & Aslin, 1990), even if they are born to congenitally deaf parents who could not have trained them in special speech patterns (Masataka, 2003). It is hypothesised that the combination of bipedalism with a lengthened developmental period born from increased brain size meant that hominid babies could no longer cling to their mothers (dated to sweat glands replacing body hair in Homo Erectus; Williams, 2011). Thus, musical mechanisms via vocal communication might have become selected at the level of infant predispositions to compensate for an inability to maintain physical contact, all the while bolstering the adaptive force of maternal bonds (Falk, 2009:69).
Others, such as Pinker (1997), suggest that music is a pleasant but unnecessary by-product of evolution, an activity that exploits human capacities (i.e. auditory scene analysis, sensitivity to pitch-patterns, motor control etc.) which have arisen for other adaptive reasons, “music appears to be a pure pleasure technology, a cocktail of recreational drugs that we ingest through the ear to stimulate a mass of pleasure circuits at once” (p.528). Such non-adaptationist accounts hold that “music has no zoological utility” (James: 1890:212), but this is an approach implicitly treating music as ethnocentrically patterned sound- commodified, technologized and alienated from the body- like that in the western world. This perception of music as nothing more than an ‘auditory cheesecake’ implies a superficial relationship to human biology, one that misses the point entirely; our distant ancestors would not have benefitted much from forming barbershop quartets, music in this culturally-constructed sense would never have evolved or become adaptive. What is relevant is to consider a natural biologically based musicality- a vehicle of self-expression through variations in pitch, rhythm, tone and timbre, like that in birdsongs, baby-babble and primate vocalisations. Recent non-adaptionist proposals have acknowledged this distinction and altered their arguments: such as music being built upon recent Theory of Mind to promote affective engagements (Livingstone & Thompson, 2009), or music’s connection to evolutionary ancient socio-emotional brain circuitry (Panksepp, 2009). These are stronger claims, but still fail to consider how music may non-adaptationally shape human brain functions (Patel, 2010).
Anatomically speaking, its antiquity can be paired to the phylogenetically older parts of the brain (e.g. cerebellum) which are activated during musical experience (Levitin, 2006), though also happen to play a significant role in linguistic processing (De Smet & Paquier, 2013). To disentangle the two, medical case studies of aphasiac patients provide an insight into their neural relationship because, if music were derivative of language, we would expect an impairment to affect both systems, or vice versa. Non-fluent aphasias generally result from lesions to the white-matter fibre tracts underlying the frontal lobe, resulting in limited and effortful verbal output, often in the face of otherwise intelligent behaviour. Strikingly, many patients can sing familiar songs with great fluency (Racette, 2006), and maintain other musical behaviours despite a chronic incapacity to talk. Most notably, 20th century composer Vissarion Shebalin suffered from several strokes which left him partially paralysed and unable to understand or produce speech. Despite such severe afflictions, Shebalin continued to analyse and write compositions, said to be equally great prior to his illness, until his unfortunate death. One could argue that professional musicians like Shebalin are untypical of the human mind because their vast dedication to music might have led to the development of extra neural networks: e.g. the left planum temporale region of the brain is much larger in musicians than non-musicians (Schlaug, 1995), and musicians who received training before puberty have better memory for spoken words (Chan, 1998). Nevertheless, an overwhelming number of documented lesions consistently report a loss of language-processing, but preservation of musical abilities, across all populations of adults (Callan, 2006; Schlaug, 2009; Norton, 2009) and children (Miller, 1989; Patel, 2010). Moreover, the converse effect of language without music is manifest as amusia, a condition marking the loss of musical abilities despite language remaining intact. Usually caused by brain degeneration (e.g. haematoma- a tumour filled with blood), patients’ perception of music becomes more like noise than sound, and their productions become frustratingly stunted, similar to the ‘tip-of-the-tongue’ phenomenon we sometimes experience with half-forgotten words. In contrast to Shebalin’s aphasia, another 20th century composer Maurice Ravel progressively suffered from amusia causing ‘blunders in writing’ and ‘puzzling performances’, until he formally retired altogether (Sergent,1993). According to his neurologist, Ravel was still able to express himself clearly and understand speech, but died acutely aware of his illness preventing him from transposing the music in his mind (Alajouanine, 1948). Other clinical cases documenting amusia strengthen the posited double dissociation between language and music (Steinke, 2001; Piccirilli, 2000; Ayotte, 2002). This is not to say they are completely independent, but should be treated as intertwined modules with developmental and evolutionary independence (see figure 8)
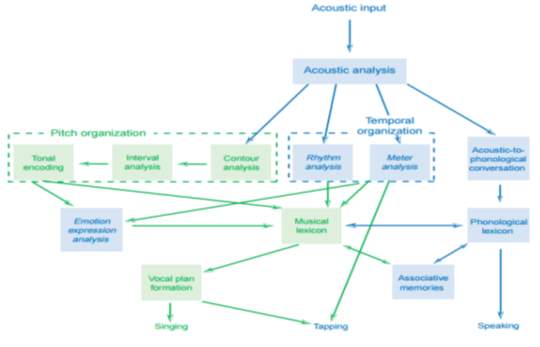
Figure 8. A modular model of music processing. Each box represents a processing component, and arrows represent pathways of information flow or communication between components. A neurological anomaly may either damage a processing box, or interfere with the flow of information between them. Component domains specific to music are green, others in blue. Taken from Peretz, 2003: 670
3.3 selective pressures for a musical system.
 Firstly, we must consider Tinbergen’s ‘four questions’ (1963). It is the order of behavioural ecology to approach any trait through this schema of complementary categories. An integrated understanding of musicality must answer all 4 questions to triumph as a complete biological explanation. (see figure 9)
Firstly, we must consider Tinbergen’s ‘four questions’ (1963). It is the order of behavioural ecology to approach any trait through this schema of complementary categories. An integrated understanding of musicality must answer all 4 questions to triumph as a complete biological explanation. (see figure 9)
Figure 9 : Tinbergen’s 4 questions, organised. Taken from Nesse 2013:681
3.3.1 Musicality: Mechanism & Ontogeny
As previously touched upon, Darwin proposed a musical protolanguage by emphasising the significance of voluntary vocalisations in his broad theory of evolution (Egnor and Hauser, 2004): “…before acquiring the power of expressing their mutual love in articulate language, [hominids] endeavoured to charm each other with musical notes and rhythm” (Darwin, 1871:880). In sum, early humans adapted by communicating in a song-like manner, and those musical displays were governed by sexual selection, analogous to how peacocks parade their plumage, giving rise to vocal learning (the ability to imitate complex vocalisations). Human infants’ remarkable acquisition of language rests on the ability to mimic vocalisations from auditory inputs (Hauser, Chomsky & Fitch, 2002), so Darwin was correct on a mechanistic level to posit vocal learning as a cornerstone requirement for human speech and song. While many animals are apt auditory learners (e.g. dog responding to sound commands), just a paucity of birds and only humans possess the cortico-motor connections between the cortex (part of the brain under volitional control) and neurones directly controlling the larynx (Carroll, 2006; Jorpens, 2002; Striedter, 2004) required for vocal learning; its absence in primates may partly explain their linguistic and musical stagnation.
It is believed that humans developed these connections via the cortico-spinal tract (major pathway connecting motor cortex to relevant neurones) and the cortico-bulbar tract (controlling the jaw, face and tongue). Developmental processes underlying axonal growth and synapse-formation between them became “exapted” to generate the synergy necessary for vocal learning (Scharff and Friederici, 2013:59). But why did this develop in humans and not in other primates? For one, Deacon (2000) proposes an explanation of bodily ontogeny involvingcompeting call-systems. Most mammals share a basic faculty for producing species-specific calls located in the periaqueductal grey (PAG) part of the midbrain (Holstege and Ehling, 1996). This ‘control centre’ produces a mammal’s entire vocal repertoire, analogous to innate vocalisations like laughter in humans, which does not rely upon cortical input. Deacon suggests that during development, there is competition between the prepotent brainstem system and the innate vocal system projections from PAG. Over many generations, competitive trophic processes sculpt these connections so that gross quantitative changes in cell numbers affect circuit organisation and ultimately behaviour via ‘Baldwinian selection’- initially learnt innate calls become unlearnt for voluntary voicing. This hypothesis has variable testable predictions, and even suggests that direct cortico-motor connections could be elicited experimentally in other mammals if the innate call system was reduced by lesions at the right stage of development (along the lines of Roe et al, 1992). While speech motor-control is closely tied to the vocal modality of communication, it is not synonymous for human language (songbirds with these capacities don’t have language). However, this aspect of speech provides a simple and clear example of how a neural circuit involved in spoken language might have evolved via exaption of a pre-existing motor circuit found in other primates.
3.3.2 Musicality: Phylogeny & Adaptive significance
When pondering why humans had protolanguage and music, Darwin reconciled an adaptive roleby imagining a musicalfunction in courtship. This framework of sexual selection would suppose one gender of humans to become disproportionally musical upon sexual maturation and the other to be better listeners, akin to the mating rituals of the dimorphic superb bird-of-paradise (Coyne, 2007). Though teenage boys often turn to electric guitars to woo their counterparts, this dimorphism is wholly incompatible with observable language patterns- linguistic and musical ability are acquired at infancy and equally across both sexes (Kramer, 2003).
The most compelling explanations are the ‘social hypotheses’ concurrently holding that language is great at conveying information, but since we cannot specify what kinds of information grammar is best adapted for, it is just as likely to have evolved to facilitate our navigation of our complex and dynamic social world. For one, the Scheherazade effect denotes how language evolved to advertise our value as prospective mates, and once mated, to service that relationship by maintaining the mate’s continued interest (Miller, 1999). Alternatively, Deacon (1997) pointed out that humans are somewhat peculiar in forming monogamous mating arrangements within large multi-male/multi-female social groups where either sex might be away for significant periods of time (Lovejoy, 1987); to minimise the risk of ‘mate-theft’, we developed social contracts and concurrent language to identify individuals who are ‘spoken for’. However, the Scheherazade hypothesis is more likely to have evolved for more other purposes, only then exploiting sexual selection, and though certain birds face the same problems Deacon identifies, they manage well without language (Emlen, 1984).
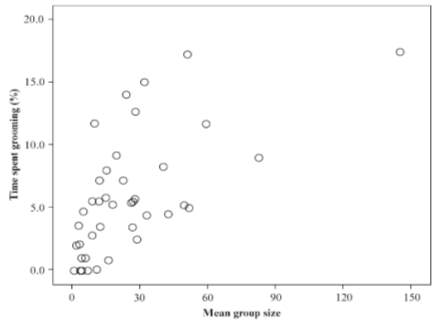
A stronger possibility is that language evolved to better service social bonds by substituting social grooming, the main mechanism used by primates to service their relationships and maintain alliances: If you scratch my back, I’ll scratch yours (Dunbar, 1993; Lehmann, 2007). For humans, as with all primates, effectively bonded social groups are essential for successful survival and reproduction, embodied in our ‘Theory of Mind’ (Astington, 1993) and more advanced forms of ‘intentional stance’ (Kinderman & Dunbar, 1998), because co-operative altruism invokes trust and obligation, a behaviour favouring the reproductive success of an organism’s relatives via Kin selection (aka, ‘Hamilton’s rule’, 1964). However, grooming time amongst primate groups makes up under 20% of the daytime (figure 10), an exogenous limit set by competing demands of core abilities (foraging, resting etc.), thus creating a glass ceiling linearly related to social group size because attending to so many peers is restrictive on one’s time budget (Dunbar, 1996). To clarify, grooming is not a purely hyginic activity. Although it does serve to clean the fur of primates, it has more in common with massages and other forms of affllative physical contact in humans like cuddling, petting etc (Dunbar, 1991), known to invoke endorphins and pleasure (Goosen, 1981).
Figure 10. Mean time devoted to social grooming by individual primate species plotted against mean group size. Taken from Lehmann et al (2007)
This constraint made it adaptive to find an alternative mechanism better suited to maximise efficiency during social bonding time. The solution, language, allowed individuals to service multiple allies and liberate both hands (amongst other advantages, see Figure 11) through low-cost emotionally-bound vocalisations. At this point, semanticity was not explicitly required, all that was necessary was to convey a message of social commitment: I consider it more important standing here talking to you than being elsewhere with anyone else. In time, enlarging group size and maximising success required more sophisticated communication; for instance, size of vocal repetoires correlate with group size and time spent grooming (McComb & Semple, 2005), thus the role of vocal exchanges as a means of supplementing grooming is reflected in increasing vocal complexity. Indeed, this correlation is well-grounded in studies measuring animal communication, even outside groups of primates, like flocks of chickadees whose uncertainty/complexity of vocal calls is greater in larger flocks than smaller ones (Freeberg, 2006).
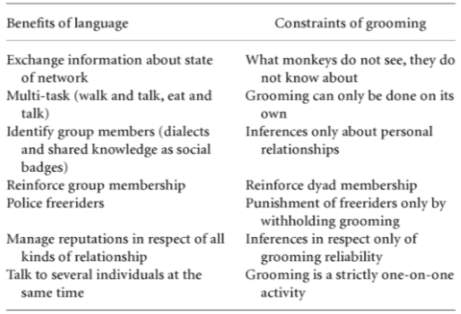
Figure 11. The mechanistic advantages of language over social grooming. Taken from Bothan & Knight (2013:10)
To picture how this might have been, comparative primatology is most useful in drawing phylogenic clues: hominid behaviours may have initially resembled that of African Geladas whose complex vocal exchanges, likened to “chorusing” (Davies & Krebs, 2012), are greatly adaptive. They engage in dyadic exchanges to maintain social contact with partners occupied by feeding etc., whilst also allowing members of a unit to monitor the surroundings without continuously breaking off. Geladas socialise in great numbers (200-300), so becoming separated from primary coalition partners would be rather troubling, thus I believe that individuals would be driven to further complexify their vocalisations in order to stand out, consistent with the semantics-first transition assumed by HPL (2.2), all the while perpetuating vocal development (3.3.2). Then, language evolved via an intermediate musical phase, because language no longer provided the psycho-pharmacological mechanism driving primates’ grooming (i.e. the production of opiate endorphins secreted to combat pain; Keverne & Martensz, 1989). If full language appeared recently (i.e. with anatomically modern humans), then there is a bonding gap between what can be done with grooming and the point at which language kicks in as a substitute. If this gap was filled by a musical type of ‘chorusing’, the endorphin-capacities of music would provide a natural bridge (Robb, 2014; Edwards, 2016), otherwise it’s unclear how hominid societies bonded between the intervening period.
Surely, a form of musical communication was operational around 500KYA according to phylogenic data. Individual hominid fossil taxas mark a significant break through the social glass-ceiling shortly after Homo Heidelbergensis (rapidly rising to 37% with Homo Sapiens; Aiello & Dunbar, 1993), which is consistent with anatomically modern forms of thoracic vertebral canals (indicative of breathing control for speech; Jungers, 2003) and hypoglossal canals (reflecting motor control over tongue and vocal space; Kay 1998). Note that the phylogenic record cannot differentiate between singing and language since the same control over breathing and vocal apparatus is needed for both. Thus, the 0.5MYA rubicon may mark the appearance of some form of intensely musical-like exchanges, with fully grammatical language emerging later perhaps at the advent of anatomically modern humans ~200KYA. As genetic bases for these different systems become better understood, there is a real possibility that this hypotheses may be confirmed by “fossils” left in the human genome allowing us to discover the sequence of selective events that drove evolutionary components to fixation, such as the late “grammar gene” (Enard, 2002; Carroll, 2006).
3.4 Possible evolutionary trajectories between music and language
As we have seen, language and music coexist through similar processing mechanisms (e.g. Peretz, 2006; Patel, 2008), yet presently perform like separate but anatomically homologous behaviours (Cross, 2006). The musicality of language, whether separate or united, may be interpreted via 3 evolutionary trajectories (Figure 12).
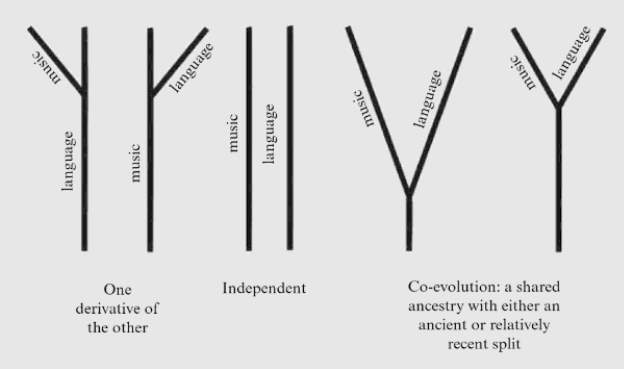 The first is a musical/linguistic precursor to protolanguage, giving rise to a system of communication or expression with the other deriving from it. This is the position that non-adaptionists like Pinker (1995) hold viewing music as a recent cultural invention occuring after the biological evolution of language.
The first is a musical/linguistic precursor to protolanguage, giving rise to a system of communication or expression with the other deriving from it. This is the position that non-adaptionists like Pinker (1995) hold viewing music as a recent cultural invention occuring after the biological evolution of language.
Figure 12. Possible evolutionary relationships between language and music, taken from Brown (2000)
Next, the “independent model” stresses the differences between music and language to have evolved as completely independent systems with any overlap as a consequence of cultural history. This is the implicit position of the majority of studies on language origins from the last 2 decades (e.g. Bickerton, 2000) that have found no need to make reference to musicality. ,intrinsic connection. Brown rejects these models as unable to explain the “deep similarities” in music and language.
Finally, a model of co-evolution posits that music and language evolved from an ancient form of communication and expression with elements of both its namesake systems, but did not truly resemble either, “the common features of these two systems are neither musical nor linguistic, but musilinguistic”. Brown attempts to explain “musilanguage” as an ancient medium characterised initially by ‘lexical tones’, a stage of discrete pitch levels able to convey complex referential meanings. Then, phrases became formed by combining lexical-tone signals into “simple, unordered phrases having higher-order meanings” (Brown, 2000:285). These meanings were referential, exapted for language, and emotional, exapted for music, ultimately bifuricating through divergence, “by which the analogous and distinct features of music and language evolved” (p.292).
Critiques of musilanguage object to the referential expressiveness of lexical tones; although the majority of the world’s languages are tonal (Fromkin, 1978), they do not use discrete level notes as music does. Phonological generativity seems implausible for a simpler protolanguage, particularly since it is a tenet of evolution that traits become more rather than less specialised with time. In fact, almost all of musilanguage is subject to criticisms of teleology since Brown’s pervasivey conflates language and speech (Fitch, 2010: 488).
Nonetheless, Brown’s diagram of co-evolution may still be relevant if specific intermediate steps in the evolution of musical protolanguage can be paired to particular hominids. This is the outlook of Paleoanthropologist Steven Mithen who draws upon anthropological and paleontological data to pinpoint steadily developing musical abilities as far back as australopithecines. His aptly named aconym “Hmmmm” denotes the cornerstones of such protolanguage: holistic, manipulative, multi-modal and musical, which became fully-fledged language once our domains of knowledge became one ‘cognitively fluid’ mentality around 200KYA. Importantly, his suggestion of a late bifurication is supported by a linear fossil record, all the while matching the pressures of kin selection laid out in 3.3.
Next, I will evaluate the ongoing narrative of musical protolanguage using Mithen’s insights to grapple the abilities of near-modern humans. The substansive issue hanging in the balance is whether the Neanderthals had archaic-like singing (if we accept that language evolved via an intermediate musical stage ~0.5MYA), or fully-fledged language (if language is unrelated or evolved independently), to reconcile the musical protolanguage hypotheses.
4. Meet the family
Briefly, Homo neanderthalensis had evolved in Europe by 250KYA and survived the bitter
where music somehow brings individuals together, thus increasing the effectiveness of group functioning
Finally, part 3 will review the musical protolanguage theory in light of the Neanderthals’ behaviour, lifestyle and cultural achievements. As a hominid species capable of some kind of communication distinguished by only marginal differences in our genus’s timespan, the comparative insights are invaluable to our understanding of anthropological linguistics, both in the name of Neanderthals and our own. As we shall see, Homo Neanderthalensis was evidently an intelligent species capable of intuitive survival in bitterly harsh living conditions. Similarly to Homo Sapiens at the time, they were able to carve complex stone artefacts, hunt big game and work co-operatively, yet they seem so different because of their stasis in technology, absence of symbolic thought and lack of language. This paradox in achievements can partly be explained by theories of evolutionary and developmental psychology, though I believe the answer may lie deeper in the kind of musical and emotional communication they developed- “a music-like communication system that was more complex and more sophisticated than that found in any of the previous species of Homo” (Mithen, 2005: p.234)
Another proposal is that musical protolanguage might have been a tool for conveying emotion, which would improve social interaction and internal decision-making (Mithen, 2009:70). Patterns of tension and resolution in music are certainly capable of eliciting an emotional response, from chills to being brought to tears. Moreover, it can be assumed that pre-modern humans displayed a range of emotions, as in the great apes who depending on species employ between fifteen and forty vocalisations tied to emotional contexts (Deacon, 1997:418). However, this innate repertoire has its human equivalence in tears, laughter, grownling, gasping, sighing and other facial and bodily responses. Indeed, ‘signals phylogenetically older than verbal language still form the most efficient channel of human interpersonal communication capable of conveying subtle feelings probably inexpressible in verbal language” (Sadowski, 2009:98). Thus it is doubtful that a musical protolanguage would arise for the purpose of something an adequate system was already in place for, a system that has remained despite the communicative power of language.
Most of the evolutionary accounts offered regarding the origin of music propose a single function, such as aiding group cohesion, sexual selection or skill acquisition depending on the type of musical protolanguage proposed.
Advancements in cognitive and social intelligence allowed our vocal imitations to pair vocalisations with objects, states, actions or emotions, and/or properties thereof, to create meanings (Holloway, 1992).
However insightful these early remarks were, it remains unclear even today how phrases became parsed, as reflected in the Compositionality vs Holophrasis debate, but compelling links between music and language must be acknowledged. At present, a renewed interest in the evolution of music, motivated by an influx of recent neuroscientific research, now renders several adaptationist accounts of how musical behaviours evolved due to their biological survival value for human ancestors. In contrast, non-adaptationist theories propose than musical behaviours are a human invention. Nevertheless, both music and language flourish so it’s essential to probe their inherent value so to resolve the chicken-and-egg paradigm upon which Musical Protolanguage hinges, “…as neither the enjoyment nor the capacity of producing musical notes are faculties of the least direct use to man in reference to his ordinary habits to life, they must be ranked among the most mysterious with which he is endowed” (Darwin, 1871: 1207).
s true since language can exist without music, suggesting a double dissociation in accounts of amusia. It is taken as a sign of both developmental and evolutionary independence.
Piccirilli 2000, Wilson & pressing, Steinke, 2001, Ayotte, 2002
Entire scientific volumes have been devoted to the evolution of music (Wallin et al, 2000; Vitouch & Ladinig, 2009), for which there are sufficient grounds to separate linguistic and musical capacities in the brain.
a mere incidental peculiarity of the nervous system” (James, 1890: 419).
3.3 Origins of music
In The Descent of Man (1871)
Medical case studies of aphasiatic patients provide the opportunity to examine the neural relationships between music and language. If one were derivative of the other, the loss of one ability ought to be an automatic consequence of the other. Alternatively, if music and language rely on entirely independent neural networks, the loss of one should have no impact on the other.
Modern music-making is apt to engage a plethora of pleasure-evoking mechanisms, and so it may prove difficult to untangle any presumed original adaptive purpose from the agglomerated mix of hedonistic mechanisms.
propose a single function (such as sexual selection). This makes sense. If music does indeed have an evolutionary origin, it would have begun by conferring a single preemitent adaptive advantage. However, as we have seen above, there is a tendency to amalgamate multiple pleasures into a single human experience. Modern musicmaking is apt to engage in a plethora in pleasyre evoking mechanisms an
Most people would accept that our brains, being part of our bodies, are also understandable as products of evolution. However, many take exception to this idea, preferring to think of the mind as being formed on individual experiences within our culture of origin. Nevertheless, the emergence of evolutionary psychology over the last decade has prompted a consideration of why humans have come to be musical. Some have proposed that musicality is a biological adaption of ancient evolutionary provenance, and thus we should show “developmental readiness for musical stimuli’ (Fitch, 2010: 506).
: musicality is a capacity we share in part with our primate relatives.
One prediction is that musicality must be a biological adaption, and thus that we should show ‘developmental readiness for musical stimuli’ (Fitch, 2010: 506), as exists for language. The adaptationist account notes that phylogenetically older parts of the brain, e.g. the cerebellum, are activated during the musical experience (Levitin, 2006). Potential adaptation values do exist for music, such as the promotion of cognitive flexibility or social cohestion. Furthermore, there is significant support for an innate musical bias in infants; Trehub (2003: 670) indicates that infants are sensitive to variations in rhythm, pitch, durational patterning and timbre, and favour IDS over speech.
This feature of music appears incompatible with Pinker’s view that music is no more than an evolutionary spin-off from language with no biological value. We don’t have emotions for free or for fun: they are critical to human thought and behaviour, and have a long evolutionary history. Our emotions are probably shared with our primate relatives and are closely related to the character of their vocalisations. Emotions are deeply entwined with the functioning of our human cognition and physiology as a control system for body and mind. It is most unlikely therefore that our deepest emotions would be so easily and profoundly stirred by music if it were no more than a recent human invention. And neither would our bodies, as they are when we automatically tap our fingers when listening to music. In fact, when we sit still, the motor areas of our brain are activated by music (Janata & Grafton, 2003)
Relationship between music and language auditory cheesecaje? Probably not because the material above indicates that music is too different from language to be adequately explained as an evolutionary spin-off. Language evolved from music, or music evolved from language? Remaining possibility single precursor shared by both but split up.
Brain evidence. Music without language
The relationship between speech and song has puzzled scholars for centuries, but it was properly elucidated by Charles Darwin who drew upon the idea of a musical protolanguage, an idea contested by early Enlightenment-thinkers like Rousseau (Thomas, 1995), by emphasising the significance of complex vocalisations in his broad theory of evolution (Egnor and Hauser, 2004): “…it appears probable that the progenitors of man, either the males or females or both sexes, before acquiring the power of expressing their mutual love in articulate language, endeavoured to charm each other with musical notes and rhythm” (Darwin, 1871:880). In sum, early humans adapted by communicating in a song-like manner, and those musical displays were governed by sexual selection, like how peacocks parade their plumage, which gave rise to vocal learning.
Advancements in cognitive and social intelligence allowed our vocal imitations to pair vocalisations with objects, states, actions or emotions, and/or properties thereof, to create meanings (Holloway, 1992).
However insightful these early remarks were, it remains unclear even today how phrases became parsed, as reflected in the Compositionality vs Holophrasis debate, but compelling links between music and language must be acknowledged. At present, a renewed interest in the evolution of music, motivated by an influx of recent neuroscientific research, now renders several adaptationist accounts of how musical behaviours evolved due to their biological survival value for human ancestors. In contrast, non-adaptationist theories propose than musical behaviours are a human invention. Nevertheless, both music and language flourish so it’s essential to probe their inherent value so to resolve the chicken-and-egg paradigm upon which Musical Protolanguage hinges, “…as neither the enjoyment nor the capacity of producing musical notes are faculties of the least direct use to man in reference to his ordinary habits to life, they must be ranked among the most mysterious with which he is endowed” (Darwin, 1871: 1207).
3.1 Music vs Language
Fitch phonology children babble ritualistic singing. Capacity for producing imitating vocalised sequences for rhythm separate from production of sound governed by semantic uses. , vocal learning. Preceding system ancestor music & language, prosodic/mimetic vocal communication. Fossil record no further back than 40,000 so have to break problem into biological record. Mechanistic level vocal learning
Similarities & differences, symbols, grammar, information transmission . IDS. Music without language. Language without music. Healing properties.
The issue is not simply the manifest nature of language and music, but the extent to which they rely upon the same computational processes in the brain
Adaptionist
Shared or independent evolutionary histories
Otto Jesperson (1895) proposed that “language began with half-musical analysed expressions for individual beings and events” (1983: 365) (claimed a holistic starting point as such that propositional meanings were drived from sung phrases. (Multi-)syllables of music phrases would have been isolated and associated with individual lexical meanings referring to different syntactic categories.
cordingly, the question remains how sentences became parsed Darwin’s insights remain astonishingly relevant despite the rapid rise in neuroscientific research on music, though it remains unclear whether parsed sentences developed out of single words or out of protomusic or a combination.
Scholars in this camp often turn to In time, musical protolanguage evolved into modern language, while modern music survived for purely entertainment purposes. My poo is so smelly, everyone in the Bragg fears it.
he points out that the development of cognition and social intelligence represents the foundation for the rise of protomusic and then to protolanguage, the likes of Bickerton (1990) and Wray (1998). The function of these initial signals might have attested to vocal imitations and by associating vocalisations with objects, states, events, actions or emotions and/or their respective properties to create meanings (Holloway, 1992). It is these vocalisations that have been the primary trigger for the evolution of language, during which the vocal tract became gradually refined allowing for a broader range of discrete utterances.
Consistent with an ewhich would have been driven by sexual selection, territorial behaviour and expressing emotions like love, jealousy and triumph. who suggested that early human
vocalizations were akin to singing rather than speech, and that musical
displays for the purpose of sexual selection gave rise to vocal learning.
as far back as the French enlightenment (Thomas, 1995), The evolutionary origins of music have absAt the turn of the 20th century, Otto Jespersen proposed that “language began with half-musical analysed expressions for individual beings and events” (1895:365). idea of a shared origin for language and music is pre-Darwinian, dating at least as far back as the French enlightenment in the 1700s (Thomas, 1995).
I believe that the musical protolanguage theory (henceforth MPL)
Evolutionary linguistics is an interdisciplinary field which consolidates evidence from
Moreover, if they were used with insufficient frequency, utterances would disappear during the vertical bottleneck stage of language transmission. In the acquisition of modern language, infants are exposed to a limited subset of the language’s total utterances with all its generative power (Chomsky, 1980), but
I believe would have been dominated by utterances descriptive of frequent and quite general events, rather than the specifics of day-to-day activity.
When learning a language, an infant is only ever exposed to a subset of the language’s total utterances (poverty of stimulus), for modern human language with its indinite generative power no child will ever hear all possible utterances. But in holistic protolanguage, relationship between utterance and meaning is not a function of its subparts and therefore unique in its place in the lexicon of similar utterances. If the lexicon is small enough then it may well be that the infant does hear all the utterances of protolanguage and transmission is learning static terms. However historical process is not perfect and replication of sounds change, thus changing utterances. Dynamic environment of hominids new situations relevant enough to warrant an increase in proto-lexicon and so situation would eventually arise where an infant no longer hears totality. Repercussions because “balance between specificity of a message” and
they would drop out of memory and lose the call. Similarly, the introduction of new phrases would be slow and difficult because it would rely on a sufficient number of individuals noticing and reproducing the form to meaning pairing
Innovation not routine. This actually happened, short of a miracle. basis of local pairwise comparison (as in Kirby, 2002
In the “smooth, gradual and beneficial” development towards hominid protolanguageA familiar scepticism towards adaptionism is the question, What good is half an eye? It goes without saying that
This gulf bridged by evolution It emerges that the Holistic account ver active field
Kirby (2002) provides a model which demonstrates that cultural transmission in a population of segmenting learners can result, under certain plausible transmission conditions, in a transition from holism to a compositionall structured linguistic system. A number of such models demonstrating this process exist (Kirby, smith & Brighton, 2004): similar results have been shown for different models of language learning (e.g. heuristic grammar inducer Kirby 2002; an associative network model Smith, Brighton, Kirby 2003). Provides proof of concept for analysis process. Modelling is robust demonstrated under assumptions of how learning works, populations structured and how meaning is constructed.
Holistic protolanguage Wray’s explanation rests upon a diachronic mechanism forming structure based on words and constraints on their combination, the accumulation of which is hard to imagine as equating to the transition to full language. For critics, segmentation is a too steep a demand for mechanism for
Wray points out the inherent weaknesses that the number of messages would be limited. Since each message must represent something different & phonetically distinct, the number of messages would be constrained by the ability of the speakers to pronouns and of the hearers to perceive sufficient phonetic distinctions. Memory capacities of early humans, similar to ours, would have imposed additional constraints. Long term limit how many could be recalled, short string length. Insufficient frequency forgotten. Introduction of new phrases would be slow and difficult.
Diachronic mechanism separate combinable words emerge. Difficult to imagine? Intricacies of eye over time
Eventually, they evolved the capacity to distinguish syllables by the segments they consist of, allowing for “…a more complex inventory of functional exchanges and thus needing a larger set of utterances” (Wray, 1998: 51)
Unfortunately, it is notoriously difficult even for speakers of modern languages to precisely define the meanings of words, and of course we cannot ask any protolanguage speakers, so discussion of the complexity of the meanings associated with hypothetical proto-forms must remain mostly conjecture
s, as a feature shared by primates and human
deliver expressions
to segment out words, leaving behind a residual template.
When a coincidental correspondence across more than one form-meaning association is found, then the learner can generalise, segmenting both form and meaning into sub-units, using analogy to create additional rules to allow their recombination. – the idea is that form/meaning correspondences occasionally occur fortuitously
Figure 6. Characteristic protolanguage utterances with their glosses. Adapted from Wray (1998)
Useful mithen 172-3, 120, 147-9, 109
What followed was the capacity to distinguish the syllables by the segments they consist of, allowing for “…a more complex inventory of functional exchanges and thus needing a larger set of utterances” (Wray, 1998: p.51)-
Eventually, the culmination of ecological pressures that produced the late Homo sapiens also produced a more complex mind that helped hominines evolve more complex phonetic communication, including syntax and morphology.. “capable of a more complex inventory of functional exchanges and thus needing a larger set of utterances” (Wray, 1998: p.51). What followed was the
For ease of expositionThe evolution to speech which is intentional, must not have been straightforward because nonhuman vocalisations are no more intentional than our cries or laughter which are innate and realised instinctively. Ape-like vocalisations must have first been co-opted for intentional use before being modified and expanded into speech
Living fossils idiomatic expressions. Look like tebima. Objections tallerman (continuity). Contra Mufwene, mithen book. Tallerman criticisms of continuity.
Mitteilungsbedürfnis’ (the urge to express and share feelings)
Wray (1998), in supporting a holistic protolanguage, appears to sidestep the issues of meaning origins by invoking continuity with primate communication. Wray argues that the vocal and gestural signals of primates function in relation to “selfpreservation, the preservation of others and the servicing of and adjustment of relationships” (1998, p.51). In this way, the argument goes, the signals are intentional, and being employed to achieve a definite effect in an interaction, and are thus propositional in content: “The signals are holistic: there are no component parts that could be recombined to create a new message” (ibid
To justify this, studies of comparative primatology point towards continuities in behaviour, perception, cognition and neurophysiology between human speech and primate vocal communication, thus positing a shared system in our deep past (Seyfarth, 2004, 2005)
words or So human language evolved from a communication system very similar to that used by primates.
The latter holistic account, protolanguage was a system in which individual signals, lacking in internal morphological structure, wholly conveyed entire complex propositions rather than semantic atoms. An utterance is holistic when it is a “complete communicative act” (Arbib, 2008: p.154) exclusively associated with one meaning encoded in the whole proposition, not the sum of its parts, and “there are no component parts that could be recombined to create a new message” (Wray, 1998: p.51)
Messages are ‘holistic’ speech-acts, centrally pragmatic in function This is not to dismiss the notion of compositionality, but rather there important stages exist important prior stages
Disentangle pursue a timeline. Insist On the other hand, another hotly debated approach is fronted by Wray (1998, 2000) as an alternative to the multi-word syntax-free intermediate stage of protolanguage (Arbib and Bickerton, 2010
it would have been unsuitable for socio-interactional expressions directed towards the physical, emotional and perceptual manipulation of the hearer (Wray, 1998;2002). It is incompatible with the subtle and sensitive communication required for the development and maintenance of social relationships, which, as I shall argue below, was critical for the evolution of vocal communication in early hominids
Wray proposes an analytic analysis to explain the emergence of compositional languages. Compositional languages do not arise through the combination of meaningful words, but rather through the correlation between features of meaningful messages and aspects of the situations that these utterances describe.
Nonetheless, a ‘words without grammar’ type of proto-language must still tell us how the words, produced phonetically, started. It is necessary to account for the transition from prephonetic words to phonetic words
From Mithen ch10,148. Subject to lots of ambiguity (ch1). Unsuitable for ‘other kinds of messages’-physical,emotional, personal. Look at Wray 1999, 2000.
Bickertonian protolanguage would have been too powerful for hominids and ealry humans. Availability of words allows novel messages to be constructed, but the majority of social communication among non-human and humans (e.g. good motning, how are you) is holistic in character- a set of routines that are understood and responded to with minimal processing. Unsuitable for development an maintenance of social relationships, and social life appears to have bee nprincipal selective pressure for evolution of vocal communication in early hominds.
whilst remain understood by one must structure must be recognised in the signal For Bickerton, the transition between these two forms of language was a biological one. Some genetic change or
changes led to a novel language faculty that enabled individual language users to go beyond their cultural heritage (a syntax-free protolanguage) and innovate a modern language. Exactly how this change in the linguistic system would progress cannot be known for sure, but we might see parallels in the process of creolisation whereby children innovate a full human language after exposure to a pidgin.
Protolanguage contains a limited set of word-like units with simple atomic meanings (bickerton 1995; Tallerman, 2007), associated with basic pre-existing cognitive concepts, effectively the ancestors of modern nouns and verbs
Cite This Work
To export a reference to this article please select a referencing stye below:
Related Services
View all


Related Content
All TagsContent relating to: "Children"
Dissertations looking at all aspects of children, their health and wellbeing, the impact of advances in technology on childhood and more.
Related Articles

DMCA / Removal Request
If you are the original writer of this dissertation and no longer wish to have your work published on the UKDiss.com website then please: