Examining Object Recognition and Object-in-Place Memory in Plateau Zokors, Eospalax Baileyi
Info: 8535 words (34 pages) Dissertation
Published: 9th Dec 2019
Tagged: Animal Sciences
Examining Object Recognition and Object-in-Place Memory in Plateau Zokors, Eospalax baileyi
Abstract
Object and Object-in-place associative (OiPA) recognition memories are important skills for the survival and fitness of the subterranean rodents due to the complicated underground circumstances and the intricacy of the burrow systems to circumvent unnecessary underground excursions which is usually associated with higher energy costs or a possible conflict with conspecifics. The two types of memory are formed during a prior experience with object and the spatial discrimination of its location which allow subsequent retrieval of previously encountered objects and their locations as well. Our study aims to examine the Object and (OiPA) recognition memory in plateau zokors (Eospalax baileyi) and test the hypothesis that their underground life exerts a sort of gender-specific differences in memory functions using Novel Object Recognition (NOR) and Object-in-Place (OiP) paradigms. Animals were tested in the NOR with short (10 min) and long-term (24hrs) retention intervals and in the OiP (30 min) retention intervals between the familiarization and testing sessions. Plateau zokors showed strong novelty preference for novel objects manifested by longer exploration time for the novel object after 10 min inter-trial interval but failed to remember the familiar object when tested suggesting a lack of memory trace for the familiar object after 24 hrs. In OiP test, zokors effectively formed an association between the objects and the place where they were encountered resulting in a higher duration of exploration to the switched objects. However, both genders showed equivalent results in exploration time during the NOR and OiP tests which eliminate the possibility for discovering gender-specific variations in memory performance. Taken together, our study illustrates robust novelty preference and an effective short-term recognition memory of plateau zokors without marked sex-specific differences.
1. Introduction
Sensory perception plays a vital role in every aspect of animal life such as food searching, communication with conspecifics (Kott et al. 2010) and avoidance of heterospecific predators which, inevitably, maximizes the animal chance for survival (Barth 2012). In the same vein, sensory perception is a key element in recognition memory where animals make extensive use of sensory modalities, such as vision (Zoccolan 2015), touch, (Diamond et al. 2008) and smell (Uchida and Mainen 2003), when exploring and interacting with objects in their environment. However, life underground is not an easy lifestyle for nearly 250 mammalian subterranean species which survive in an extensive underground burrow system separated from the surrounding environment by mounds of soil (Bennett and Faulkes 2000). Nevertheless, these burrows, although protecting animals from predation and climatic changes, deprived of light and ventilation which consequently tackle visual, olfactory and auditory cues available aboveground (Bednářová et al. 2013) and might hinder the cognitive ability of the subterranean dwellers to be aware of and extract meanings from sensory inputs.
In wild niche, exploration of novel objects or places that depart from normal circumstances comprising ambulatory and investigatory responses which help acquainting animals with surroundings, secure better information, avoid danger, and gain prospective knowledge, which can affect their fitness and survival as well (Lee et al. 2016). These exploratory responses, if not deployed wisely, may consume much time and energy that could be used for alternative activities (Winkler and Leisler 1995). However, a popular claim may contradict the previous concept as wild rodents demonstrate significantly higher neophobic [avoidance of novel items in familiar surroundings (Barnett 1958)] responses to unfamiliar objects in their habitats Calhoun 1963, Mitchell D 1976). This novelty preference phenomena had not been yet extensively studied in subterranean species. Furthermore, memory studies in subterranean rodents are not sufficient and had been curtailed on examination of spatial orientation and memory (Kimchi and Terke 2001, Kimchi and Terkel 2003, du Toit 2012, Lovy et al. 2013) and only few studies (Burda et al. 1995) have been conducted to assess recognition memory in subterranean species.
The pioneer work of Ennaceur and Delacour (1988) and others had emerged into new recognition memory paradigms for rodents known as novel object recognition (NOR) and object-in-place (OiP) tests. The standard procedures are very straightforward, the animal is first exposed to a sample object (Two or more identical objects in specified locations) and after a retention interval, one object is either substituted with a novel one (NOR) or two objects switched to a novel position (OiP) (Antunes and Biala 2012). The interval of the retention phase could be varied from minutes to measure short term memory or extended to over 24 hours for long term memory examination. The principals of the tests depend on the natural tendency of the rodents to explore novel objects or novel locations in the presence of familiar one and the variations in the testing modalities has been used to investigate types of recognition memory in rodents (Leger et al. 2013). NOR and OiP have gained extra privilege and popularity above other memory tests due to lack of necessity to apply any emotionally aversive stimuli such as food or water restrictions and the convenience to obtain relevant behavioral measurements (Blaser and Heyser 2015). Several studies have documented the importance of different sensory information from the environment as visual, olfactory, and somatosensory stimuli, in object recognition task, where impairment of environmental cues disturbed the ability of the normal rats to discriminate the novel object in darkness during the testing phase (Albasser et al. 2011). Novel object/Location recognition memory has been investigated in many different mammalian species including mice (Wang et al. 2017), rats (Arfa-Fatollahkhani et al. 2017), hamsters (Muller et al., 2015), monkeys (Melamed et al., 2017) and Gottinger minipigs (Sondergaard et al., 2012).
Plateau zokors (Eospalax baileyi) are a typical subterranean rodent species inhabit the Qinghai-Tibetan Plateau (QTP) (Su et al. 2015) where they live and forage in very complicated burrow systems with blind endings (Zhou and Dou 1990) representing a dark ecotope with limited cues for spatial and cognition tasks (Wegner et al. 2006). Their feeding and excavating activities severely destroying the grasslands in the QTP (Kang et al. 2007). Although they may show infrequent aboveground trips (Norris et al. 2004), most individuals remain their entire life underground surrounded by permeant darkness. During winter, harsh environmental conditions prevails at the QTP with shortage of food resources mandates zokors to hoard their food in specialized nets to avoid possible food shortage during winter (Zhang 2007). These limitations in food resources should thus pose a strong selective pressure in plateau zokors for effective recognition memory which may facilitate their spatial orientation and avoidance of energy-consuming exploratory and digging activities. Interestingly, plateau zokors exhibited sexually dimorphic variations in various aspects of life. For example, both sexes dig extensive burrows for foraging and food hoarding, but females live in burrows lie on deeper distances (2-2.5 m depth) than burrows of males (0.8-1.5 m) (Shao et al. 2015). Furthermore, males keep wider home ranges (1790±720) m2 compared to the smaller female range (260 ± 112) m2(Zhang, Fan & Zhou 1993). These sex-specific differences in home-range size may reflect different cognitive abilities between genders which make this point deserve further investigation. Here, we used the NOR and OiP tasks to test the recognition abilities and novelty preferences in males and females plateau zokors. Despite the extensive research on this topic, to the best of our knowledge this is the first study examining the novel object and location recognition memory in plateau zokors and the possible differences between genders. Our study provides novel understandings into the dynamics of recognition memory formation in the plateau zokors.
2. Materials & Methods
2.1 Animals
All procedures were approved by the Institutional Animal Care and Use Committee of the college of grassland sciences of Gansu Agricultural University. Our experiments were conducted in the Laboratory of Animal Behavior, College of Grassland Sciences, Gansu Agricultural University, Lanzhou, Gansu Province, China. The plateau zokors (Eospalax baileyi) inhabits the Qinghai- Tibetan plateau, Gansu Province, China. Wild zokors were captured alive in a specialized tube traps (Baoji Ludi xincheng Co., Ltd, Xian, China) from the grassland fields of Tianzhou Tibetan Autonomous County northwest part of Qinghai- Tibetan plateau (37.168346°N, 102.811201°S), Gansu Province. No trap mortalities or injuries were recorded. Animals were then transported to animal room and upon arrival, their health status was checked, weighed, sexed and individually housed in plastic-bottomed wire cages (45 × 30 × 20 cm). Animals were fed ad libitum with carrots, potatoes, sweet potatoes, cabbages, and a commercial rodent diet pellets supplied by Shenyang Maohua Science and Technology Co., Ltd, Liaoning, China. Wood shavings (Pet-Link®, Pet-Link Co., Ltd, Guangzhou, China) were used as bedding material and changed every 2 weeks or when necessary. Cages were washed every 4 weeks. Animals were kept on a reversed 12-h light/dark cycle at 23±1°C and humidity 55 ± 5% in an animal room at Gansu Agricultural University. Animals were accustomed to daily procedures including handling, weighing, food delivery and bedding change for at least two weeks before starting the experimental procedures.
2.2 Apparatus and surrounding environment
The testing apparatus was a plastic, light blue rectangular arena, measuring 70 cm x 50 cm x 40 cm high with a solid floor. The apparatus was situated in a quiet separate room, surrounded by a black curtain with an infra-red video network camera (Model. ZKXC.TD136U2RZT, Zhongke Electric Co., Ltd, Shenzhen, China) mounted above the apparatus and connected to a monitor and a computer in an adjacent room for recording and monitoring animal behavior. All tests took place between 9.00 am to 13.00 pm which represents one of the two daily peaks of activities in plateau zokors according to the light-dark cycle (Zhang 2007). Animals were tested in a complete darkness conditions to mimic the underground niche.
2.3 Procedures
All animals were experimentally-naïve to the testing conditions and have not undergone any behavioral experiments before. Every animal was firstly habituated to the testing arena by placing them the day before testing in the empty apparatus for a 10-min habituation session. After receiving the habituation session in the empty arena, animals were returned to their home cages and the arena was cleaned with 50% ethanol after removing urine and dirt and left to dry. Animals were always handled and transferred from the animal room to the testing room by a familiar person thereby minimizing stress and improving the validity of the results.
A. Novel Object recognition task (NOR)
This experiment examined both short-term and long-term memory in males and females plateau zokors. Twenty-six (14 males and 12 females) and twenty-two (12 males and 10 females) subjects were used for short-term and long-term memory testing, respectively. One day after habituation, two identical objects (2 plastic bottles, Fig. 1-A) were placed in the arena, fixed to the floor with double-face strong adhesive tape, 20 cm apart and 10 cm from the wall for object familiarization session (Trial 1). The subject was placed in the empty place between the two objects facing away from objects and allowed five minutes to explore them. After exposure to the identical objects, the animals were returned to their home cage and the arena was cleaned as before. After a 10-min (Short-term) or 24-h (long-term) inter-trial interval, zokors were released into a similar arena with one of the familiar objects was replaced with a novel one and let to explore the familiar and novel objects during a 5-min test phase (Trial 2). Both familiarization and test phases were recorded and later analyzed using computer software. The position of the novel object was counterbalanced among zokors to preclude side biases. Object-in-Place task (OiP)
A. Object-in Place task (PiO)
This experiment assesses spatial recognition memory and discrimination skills in males and females plateau zokors. Twenty-One (13 males and 8 females) individuals were used in this experiment. Animals were habituated to the empty arena as described before. One day later, animals were allowed to explore the arena that contains four different objects [Cylinder (Object 1), Rectangle (Object 2), Semi-hemisphere (Object 3) and Pyramid (Object 4), Fig. 1- B] in the corners 10 cm from the walls for a 5-min familiarization session (Trial 1) and then returned back to their home cages. The arena and objects were then cleaned as before and two of the objects (Object 2 and Object 4) were switched in position while Object 1 & 3 remined in their original locations without any change. After a 30-min retention phase, animals were sent back to the apparatus and allowed to explore the four objects for a 5-min test phase (Trial 2). Both familiarization and test phases were recorded and later analyzed. The time spent investigating the four objects in the in the familiarization phase was compared with the time spent investigating the four objects in test phase. Object position was counterbalanced between animals. Intact object-in-place memory occurs when zokors spends more time investigating the 2 objects in different than the 2 objects in the same locations.
2.4. Behavioral Measurements and Data Analyses
All recordings were analyzed using the Behavioral Observation Research Interactive Software (BORIS, v. 2.95, University of Torino, Torino, Italy). Behavioral measures collected during familiarization and test sessions comprised the amount of time for locomotion and the amount of contact with each object. Object contact was only started when the nose come in contact with the object which discarded certain unwanted behaviors such as object climbing, side contact, tail contact and resting next to the object. Any animal that didn’t achieve at least 5 sec contact to any of the object during the familiarization session and 1 sec during the test session was excluded from the analysis. This protocol was followed according to (Cyrenne D.L. & Brown 2011)
Side Preference Index for object recognition task was calculated as the duration of time spent interacting with the right vs left object in familiarization session as follows: [(Time with Right – Time with Left)/ (Time with Right + Time with Left) x 100]. Positive values denote preference for right object and negative values indicate preference for the left one and 0 scores indicate preference for neither objects. In the same vein, Preference for Novelty Index was also calculated for test phase: [(Time with Novel- Time with Familiar)/ (Time with Novel + Time with Familiar)]. Positive values represent preference for novel object and negative values indicate preference for the familiar one (Antunes and Biala 2012). Moreover, the place discrimination index was calculated by using the equation: [the time spent with the objects (Combined for objects 2 and 4) moved to a novel places/the total time spent in exploring both the objects moved to a novel place and the objects remaining in the familiar place].
2.5. Statistical Analyses
SPSS 22.00 software was used for all analyses. Repeated-measures analyses of variance (ANOVAs) were used to test gender differences in locomotor and object contact measures across the two trials or within the same trial. Student’s t-test was also used to examine the difference in side preference, Preference for novelty and place discrimination indices values between trials (2-tailed). Duncan’s Post hoc multiple comparison test was conducted where appropriate. The level of significance at which the null hypothesis was rejected was α = 0.05.
3. Results
3.1. Object recognition short-term memory
Using a 2 × 2 repeated measures ANOVA of amount of time spent locomoting inside the arena, with sex as the between-subjects factor and trials (Trial1 vs. Trial2) with 10 minutes inter-trial interval as the repeated within-subject factor, there were no significant main effects of sex (F1,24=2.18, p=0.15), trial (F1,24=2.96, p=0.10) nor significant interactions between sex and trial (F1,24=0.25, p=0.62). Plateau zokors spent 78.89±6.67 and 69.02±5.10 sec moving inside the arena in Trial 1 and Trial 2, respectively. The total amount of exploration time didn’t significantly change (F1,24=0.19, p=0.67) across trials (33.00±3.25 and 31.57±3.79 s for Trials 1 & 2, respectively) and there was no main effect of sex (F1,24=0.82, p=0.37) or Sex x Trial interaction (F1,24=0.07, p=0.80).
Figure (2-A) shows the preference for the both objects exhibited by the plateau zokors during familiarization session. Plateau zokors didn’t show preference toward any of the objects in the testing arena as repeated-measures ANOVA with sex as the between-subjects factor and side (Right vs. Left) as the within-subjects repeated factor revealed that neither sex (F1,24=0.53, p=0.47), side (F1,24 =0.03, p=0.86) nor Sex x Side interaction (F1,24 =2.25, p=0.15) had any significant effect on the mean duration of time spent by zokors exploring both right and left objects [Right (16.40±2.36 s), Left (16.61±2.59 s)] resulted in a side preference index of 0.005±0.11 which eliminate the bias toward either right or left objects. On the contrary during the test phase (Fig. 2-B), changing one of the familiar objects with a novel induced significant side preference (F1,24=8.06, p=0.00) in plateau zokors to explore novel object (19.39±2.51s) more than the familiar one (12.18±2.01s). However, no significant effects of sex (F1,24 = 0.73, p=0.40) or Sex x Object interaction (F1,24 =0.10, p=0.76) were obtained from the analysis. For preference of novelty, both males and females showed positive means toward novel objects (0.27±13.52 and 0.24±10.25, respectively) but t-test revealed no significant difference between genders (t23=−0.15, p=0.88). When data of both genders were combined, t-test also showed that the means of novelty preference between trials (Fig.5) significantly changed toward exploring the novel object (t49=−2.05, p=0.045) (Trial 1= -0.005±0.11, Trial 2= 0.25±0.10).
3.2. Object recognition long-term memory
Using a 2 × 2 repeated measures ANOVA of amount of time spent locomoting inside the arena, with sex as the between-subjects factor and trials (Trial1 vs. Trial2) with 24 hours inter-trial interval as the repeated within-subject factor, there was no significant main effect of sex (F1,20 =0.11, p=0. 74), trial (F1,20=2.21, p=0.15) nor were there significant interactions between sex and trial (F1,20 =2.37, p = 0.14). However, the total contact time showed significant decrease between trials (F1,20=5.79, p=0.03) as zokors spent more time exploring both objects in Trial 1 (34.00±3.65) than Trial 2 (27.88±3.71) but no main effect of sex (F1,20=0.28, p=0.60) nor their interaction (F1,20=0.95, p=0.34) was found. The exploration time of the two identical objects showed non-significant differences as repeated measures ANOVA with sex as the between-subjects factor and side (Right vs. Left) as the within-subjects repeated factor revealed that there was no significant side preference for right (10.60±2.29) or left objects (11.62±1.90) in both genders [sex (F1,20 =0.04, p=0.85), side (F1,20 =0.14, p=0.71)] nor Sex x Side interaction (F1,20 =0.06, p=0.81). Side preference was (-0.03±0.11). Similarly, during testing phase, the 2 × 2 repeated measures ANOVA showed that neither object type [(Familiar vs. Novel) (F1,20=0.08, p=0.78)], sex (F1,20=0.93, p=0.35) nor Sex x Object interaction (F1,20=0.19, p=0.67) significantly change the duration of exploration time for familiar and novel objects during the testing phase following the 24-h retention session (Fig. 3- A&B).
3.2. Object-in-Place memory
Comparison of the locomotion levels between the familiarization and test phases was conducted using 2 x 2 repeated measures ANOVA with Trials (Trial 1 vs Trial 2) as within subject factor and gender (Males vs Females) as between subject factor. The ANOVA showed no significant main effects or interactions [ Trial= F1,19=0.23, p=0.64, Sex= F1,19=0.03, p=0.88, Trial x Sex= F1,19=1.95, p=0.18] confirming that there was no statistical difference in locomotion levels between Trial 1 (56.82±4.70) and Trial 2 (56.25±4.94). However, plateau zokors spent significant more time (F1,19=7.26, p=0.01) exploring both objects during Trial 1 (20.89±3.12) more than Trial 2 (16.87±2.20) and Trial x Sex interaction was also significant (Trial x Sex= F1,19=10.44, p=0.004) but the main sex effect didn’t affect the total exploration time (Sex= F1,19=1.97, p=0.18). Further analysis by Duncan’s multiple comparison post hoc test showed that female zokors displayed the highest exploration time during Trial 1 (29.26±6.65). During familiarization session, repeated measure ANOVA revealed that plateau zokors showed no preference toward any of the four objects (Fig. 4-A) in the testing arena (Object= F1,19=0.40, p=0.63, Object x Sex= F1,19=0.76, p=0.45). However, a significant main effect of sex (Sex= F1,19=5.41, p=0.03) was found which influences the time spent exploring objects during familiarization session where zokor females spent more time exploring objects (7.31±1.14) than males (3.93±0.90). To calculate the place discrimination index, data of Objects 1 & 3 and Objects 2 & 4 (which were later exchanged in position) were combined. A place discrimination index of (-0.02±0.07) indicated that there were no preference biases to objects. However, plateau zokors showed significantly more tendency to explore the object moved to a novel place (F1,19=3.47, p=0.045) than those remained in the old locations (Fig. 4-B). No significant main effect of sex (F1,19=0.00, p=0.99) nor Object x Sex interaction (F1,19=0.28, p=0.74) was found. In order to calculated place discrimination index, exploration time for objects moved to novel places and those remained in familiar places were combined. Student’s t-test showed a significant difference (t40=3.28, p=0.002) in place discrimination index (Fig. 5) between familiarization and test trials (-1.78±6.54 and 0.30±0.07, respectively). T-test also showed that there is no significance difference in place discrimination index (t19=-0.58, p=0.57) between males (0.27 ±0.09) and females (0.35±0.13).
4. Discussion
This study examined recognition memory in plateau zokors using the novel object recognition and object-in-place tests as a model test of spontaneous recognition memory without prior training. The first experiment illustrated that plateau zokors exhibit short-term recognition memory as shown by a strong novelty preference following 10-min delay interval, but plateau zokors showed impaired long-term memory recognition for novel object after 24 hrs retention time. In the second experiment, plateau zokors can effectively locate familiar objects in novel positions after 30 minutes interval between familiarization and test session. However, our study found no general significant variations between the performances of the sexes in the two tasks.
Our results showed that zokors exhibited robust novelty discrimination during test phase after 10 minutes retention interval which reflecting a very rich recognition memory of the familiar object. Previous work has shown that rodents actively displayed preference for novel objects in the presence of familiar one following short-term retention time (Hale and Good 2005, Ennaceur 2010, Hoffman and Basurto 2013, Zhang et al. 2015, Hauser et al. 2017). Although the previous studies were conducted on the aboveground rodents, but the similarities with our results may suppose an equivalent degree of short-term memory processing and consolidation between the two taxa. Zokors are daily confronted during underground exploration and digging with numbers of objects. Therefore, recognition memory, the ability to recognize formerly encountered objects or locations (Eichenbaum et al. 2007), is imperative for the subterranean species to remember the landmarks in their underground environment which might facilitate quick identification and avoiding unnecessary exploratory endeavors that may result in excessive digging activities and increased energy expenditure (Costanzo et al. 2009). However, contrary to other wild counterparts (Cowan 1977, Kiyokawa et al. 2017), plateau zokors showed neophilic responses similar to those observed in animals live under laboratory conditions. This is usually clarified by the nature of the underground habitat where nearly lack of predation and more environmental stability prevail. The absence of changeable environmental pressures that probably inhibits novelty seeking in other wild rodents, may increase the stimulus-approach pattern observed in plateau zokors (Stryjek et al. 2012). Similarly, our results illustrated an excellent ability of zokors to link between objects and the location in which they were formerly visited. Several studies have highlighted the spatial learning and memory skills in plateau zokors (Kimchi and Terkel 2001, Schleich 2010, Mastrangelo et al. 2010). However, to the best of our knowledge, this is the first study that examined the association between recognition and spatial memory performance in subterranean rodents and plateau zokors using (PiO) task. This short-term object/location associative memory is particularly interesting in subterranean rodents because it denotes a quick acquisition of information and a comprehensive combination between recognition and spatial memory. Successful foraging, reproduction, predator evasion and territorial defense all rely on precise spatial/location recognition for spatial localization of food sources, mates and escape paths (Kimchi and Terkel 2001) to avoid unnecessary energy expenditure and/or possible conflict with conspecifics.
Obviously, there is a distinctive absence of novelty preference detected after 24 hrs in plateau zokors which is thought to be due to a lack of maintain a clear memory of the familiar object over this time. Nevertheless, there are discrepancies in across literature concerning the outcomes of long-term memory (24 h) in rodents: as several studies have shown a lack of significant retention in a long-term memory NOR task (Ennaceur and Delacour, 1988; Okuda et al., 2004; Roncarati et al., 2009; Tellez et al., 2010), a few studies have revealed the existence of long-term retention effects (Stefanko et al. 2009, Dellapolla et al. 2017). This inconsistent between different studies may arise from differences in methodological approaches and manipulation like administration of memory enhancers or suppressors, the degree of similarities between objects, the handling of animals or the level of stress in their environment. Furthermore, an elegant study conducted by Khani and Rainer (2012) on tree shrew (Tupaia belangeri) found that single familiarization session resulted in poor recognition memory to the familiar object while 3 repetitive familiarization sessions yielded significant preference for the novel object after 24 h retention interval. This might suggest that long-term recognition memory require multiple exposures to the familiar object for better memory processing and consolidation in wild counterparts. Another confounding factor that should be taken into consideration is the effect of chronic housing conditions and limited space offered to wild animals brought into laboratory. In our study, although individuals didn’t show significant signs of distress during NOR or PiO testing such as excessive bouts of inactivity or others (Bevins and Besheer 2006), but we can’t ignore the effect of the stressful housing conditions on wild zokors. In nature, zokors are free-roaming individuals that occupy a large home range of 1790 ± 720 m2 for males and 260 ± 112 m2 for females (Zhang 2007). However, under laboratory conditions animal are restricted to a very narrow home range which inevitably curtail their active movements and impose a sort of chronic stress upon them. Consequently, chronic stress is infamous of increasing glucocorticoid levels and consequently induces aberrant synaptic plasticity which can provoke deleterious effects on long-term cognitive functions and recognition memory (Elizalde et al. 2008). The effect of chronic stress on recognition memory in NOR was documented in (Park et al. 2015)where exposure to chronic stress alters the performance of stressed subjects in the long-term NOR. On the contrary, our results of the short-term NOR (10 min) and OiP (30 min) tests are most likely to be unaffected by the chronic stress resulted from the limited housing conditions which may highlight the effect of chronic stress on long-term memory consolidation and retention while in short-term retention sessions the memory of the familiar object/locations were still vivid. A study by Beck and Luine (1999) have also reported that chronic stress impairs object-recognition memory when the retention time was extended more than 1 h which supports our assumption that the impairment of long-term performance was due to a chronic stressful situation rather than a limited innate ability to remember the familiar object. Extensive research should be conducted to unravel the consequences of limited housing conditions on neural and hormonal profile of plateau zokors.
Another interesting finding in our study is the lack of gender specific differences in both short and long-term recognition memory and the object/place associative memory amongst the tested zokors. This contradicts our previous presumption of gender-specific differences for the plateau zokors although sex-specific variations in recognition memory abilities have been reported before (Sutcliffe et al. 2007, Cyrenne and Brown 2011, Bettis and Jacobs 2013, Bath et al. 2017). However, all the previous studies that demonstrated the gender differences in recognition abilities have been conducted on the surface-dwelling rodents and none of them was conducted on subterranean species.Yet, our results are parallel with similar studies on subterranean rodent that exhibited similar gender performances during memory tasks (Kimchi and Terkel 2001, Mastrangelo et al. 2010). To explain our results, the relationship between home range size and mating system has been anticipated that sexually dimorphic home-range size will trigger differences in recognition memory abilities that males have larger home ranges than do females as is the case in promiscuous species including plateau zokors. However, these home range sizes are only limited to the breeding season for mate search and during the non-breeding season both sexes displayed similar territory size (males 560 ± 101 m2 and females 640 ± 130 m2) which may explain the similarities in recognition abilities between genders in our study and in similar research work. In species with sexually isometric home range sizes no sex differences in memory skills were found (Gaulin and Fitzgerald 1989). Generally speaking, the high energetic costs of digging and the similarities of the underground environment could restrain the development of superior recognition abilities of one gender over the other, yet enforce some degree of behavioral sex differences in males. For example, during the breeding season males and females never live together in the same tunnel but instead, males start to dig longitudinal tunnels with two-branches end to intercept the female burrows and increase the chances to trap the females (Zhang 2007). Colloquially, the lack of superior recognition memory to locate the female burrows in males is compensated by some advanced digging techniques the maximize the chances to locate mates.
Finally, our results also showed that there were non-significant differences in ambulation time inside the arena between familiarization and testing sessions. The non-significant differences in locomotor activities in both NOR and OiP task may be due to the long interval between habituation and familiarization sessions (24 hrs). Also, the presence of novel objects or novel locations may provoke exploratory activities in plateau zokors directed toward exploring the arena itself.
In conclusion, our findings suggest that plateau zokors showed strong recognition memory in the NOR after short term (10 min), but not long-term (24hrs), exposure session results in appearance of novelty preference responses toward the novel object. Moreover, zokors can effectively discriminate the changes in their environment upon relocation of familiar objects in novel places. Our results also support the notion that both genders of the subterranean species displayed equal memory skills as sex differences was absent in NOR and OiP tasks. Finally, our results also highlight the need for further investigations on the housing conditions for subterranean rodents to ensure better experimental results.
5. Acknowledgement
Funding for this work was supported by the National Natural Science Foundation of China (No. 31460566); Gansu Provincial Natural Science Foundation for Distinguished Young Scholars of China (No. 1606RJDA314), the China Postdoctoral Science Foundation (Nos. 2015M572614 and 2016T90958) and Talented Young Scientists fellowship (TYSP) of the Ministry of Science and Technology of China. The authors declare that they have no competing interests. Plateau zokors were trapped by professional members of our research team. Experimental procedures were approved by the animal ethics committee of Gansu Agricultural University as well as a permission from the local authorities was required for our study.
6. References
Albasser MM, Amin E, Iordanova MD, Brown MW, Pearce JM, Aggleton JP (2011) Separate but interacting recognition memory systems for different senses: the role of the rat perirhinal cortex Learn Mem 18:435-443 doi:10.1101/lm.2132911
Antunes M, Biala G (2012) The novel object recognition memory: neurobiology, test procedure, and its modifications Cognitive processing 13:93-110 doi:10.1007/s10339-011-0430-z
Arfa-Fatollahkhani P et al. (2017) The Effect of Luteinizing Hormone Reducing Agent on Anxiety and Novel Object Recognition Memory in Gonadectomized Rats Basic and clinical neuroscience 8:113-119 doi:10.18869/nirp.bcn.8.2.113
Barnett SA (1958) Experiments on ‘neophobia’ in wild and laboratory rats. Brit J Psychol 49: 195–201.SA Barnett1958Experiments on ‘neophobia’ in wild and laboratory rats.Brit J Psychol49195201
Barth FG (2012) Sensory perception: Adaptation to lifestyle and habitat. In: Barth FG, Giampieri-Deutsch P, Klein H-D (eds) Sensory Perception: Mind and Matter. Springer Vienna, Vienna, pp 89-107. doi:10.1007/978-3-211-99751-2_6
Bath KG et al. (2017) Early life stress leads to developmental and sex selective effects on performance in a novel object placement task Neurobiology of stress 7:57-67 doi:10.1016/j.ynstr.2017.04.001
Beck KD, Luine VN (1999) Food deprivation modulates chronic stress effects on object recognition in male rats: role of monoamines and amino acids Brain Res 830:56-71
Bednářová R, Hrouzková-Knotková E, Burda H, Sedláček F, Šumbera R (2013) Vocalizations of the giant mole-rat (Fukomys mechowii), a subterranean rodent with the richest vocal repertoire Bioacoustics 22:87-107 doi:10.1080/09524622.2012.712749
Bennett NC, Faulkes CG (2000) African mole-rats: ecology and eusociality. Cambridge: Cambridge University Press. 287 p.NC
Bettis TJ, Jacobs LF (2013) Sex differences in memory for landmark arrays in C57BL/J6 mice Animal cognition 16:873-882 doi:10.1007/s10071-013-0619-x
Blaser R, Heyser C (2015) Spontaneous object recognition: a promising approach to the comparative study of memory Front Behav Neurosci 9:183 doi:10.3389/fnbeh.2015.00183
Burda H (1995) Individual recognition and incest avoidance in eusocial common mole-rats rather than reproductive suppression by parents Experientia 51:411-413 doi:10.1007/bf01928906
Calhoun JB (1963) The Ecology and Sociology of the Norway Rat. Bethesda, MD: U.S. Department of Health, Education and Welfare. JB Calhoun1963The Ecology and Sociology of the Norway Rat.Bethesda, MD: U.S. Department of Health, Education and Welfare
Costanzo MS, Bennett NC, Lutermann H (2009) Spatial learning and memory in African mole-rats: The role of sociality and sex Physiol Behav 96:128-134 doi:http://dx.doi.org/10.1016/j.physbeh.2008.09.008
Cowan PE (1977) Neophobia and neophilia: New-object and new-place reactions of three Rattus species Journal of comparative and physiological psychology 91:63-71 doi:10.1037/h0077297
Cowan PE (1977) Neophobia and neophilia: new-object and new-place reactions of three Rattus species. J Comp Physiol Psych 91: 63–71.PE
Cyrenne DL, Brown GR (2011) Ontogeny of sex differences in response to novel objects from adolescence to adulthood in lister-hooded rats Dev Psychobiol 53:670-676 doi:10.1002/dev.20542
Dellapolla A et al. (2017) Long days enhance recognition memory and increase insulin-like growth factor 2 in the hippocampus Scientific reports 7:3925 doi:10.1038/s41598-017-03896-2
Diamond ME, von Heimendahl M, Arabzadeh E (2008) Whisker-mediated texture discrimination PLoS biology 6:e220 doi:10.1371/journal.pbio.0060220
du Toit L, Bennett NC, Nickless A, Whiting MJ (2012) Influence of spatial environment on maze learning in an African mole-rat Animal cognition 15:797-806 doi:10.1007/s10071-012-0503-0
Eichenbaum H, Yonelinas AP, Ranganath C (2007) The medial temporal lobe and recognition memory Annual review of neuroscience 30:123-152 doi:10.1146/annurev.neuro.30.051606.094328
Elizalde N, Gil-Bea FJ, Ramirez MJ, Aisa B, Lasheras B, Del Rio J, Tordera RM (2008) Long-lasting behavioral effects and recognition memory deficit induced by chronic mild stress in mice: effect of antidepressant treatment Psychopharmacology 199:1-14 doi:10.1007/s00213-007-1035-1
Ennaceur A (2010) One-trial object recognition in rats and mice: methodological and theoretical issues Behavioural brain research 215:244-254 doi:10.1016/j.bbr.2009.12.036
Gaulin SJC, Fitzgerald RW (1989) Sexual selection for spatial-learning ability Anim Behav 37:322-331 doi:http://dx.doi.org/10.1016/0003-3472(89)90121-8
Hale G, Good M (2005) Impaired visuospatial recognition memory but normal object novelty detection and relative familiarity judgments in adult mice expressing the APPswe Alzheimer’s disease mutation Behavioral neuroscience 119:884-891 doi:10.1037/0735-7044.119.4.884
Hauser MJ, Isbrandt D, Roeper J (2017) Disturbances of novel object exploration and recognition in a chronic ketamine mouse model of schizophrenia Behavioural brain research 332:316-326 doi:10.1016/j.bbr.2017.06.013
Hoffman KL, Basurto E (2013) One-trial object recognition memory in the domestic rabbit (Oryctolagus cuniculus) is disrupted by NMDA receptor antagonists Behavioural brain research 250:62-73 doi:10.1016/j.bbr.2013.04.049
Kimchi T, Terkel J (2001) Spatial learning and memory in the blind mole-rat in comparison with the laboratory rat and Levant vole Anim Behav 61:171-180 doi:10.1006/anbe.2000.1565
Kimchi T, Terkel J (2003) Mole rats ( Spalax ehrenbergi) select bypass burrowing strategies in accordance with obstacle size Die Naturwissenschaften 90:36-39 doi:10.1007/s00114-002-0383-2
Kimchi T, Terkel J (2004) Comparison of the role of somatosensory stimuli in maze learning in a blind subterranean rodent and a sighted surface-dwelling rodent Behavioural brain research 153:389-395 doi:10.1016/j.bbr.2003.12.015
Kiyokawa Y et al. (2017) Two strains of roof rats as effective models for assessing new-object reaction Journal of Veterinary Medical Science 79:702-708 doi:10.1292/jvms.17-0002
Kott O, Sumbera R, Nemec P (2010) Light perception in two strictly subterranean rodents: life in the dark or blue? PloS one 5:e11810 doi:10.1371/journal.pone.0011810
Lee YF, Kuo YM, Chu WC (2016) Energy state affects exploratory behavior of tree sparrows in a group context under differential food-patch distributions Front Zool 13:48 doi:10.1186/s12983-016-0180-y
Leger M, Quiedeville A, Bouet V, Haelewyn B, Boulouard M, Schumann-Bard P, Freret T (2013) Object recognition test in mice Nature protocols 8:2531-2537 doi:10.1038/nprot.2013.155
Lovy M, Skliba J, Sumbera R (2013) Spatial and temporal activity patterns of the free-living giant mole-rat (Fukomys mechowii), the largest social bathyergid PloS one 8:e55357 doi:10.1371/journal.pone.0055357
Mastrangelo ME, Schleich CE, Zenuto RR (2010) Spatial learning abilities in males and females of the subterranean rodent Ctenomys talarum Ethology Ecology & Evolution 22:101-108 doi:10.1080/03949370903516156
Mastrangelo ME, Schleich CE, Zenuto RR (2010) Spatial learning abilities in males and females of the subterranean rodent Ctenomys talarum Ethology Ecology & Evolution 22:101-108 doi:10.1080/03949370903516156
Melamed JL, de Jesus FM, Maior RS, Barros M (2017) Scopolamine Induces Deficits in Spontaneous Object-Location Recognition and Fear-Learning in Marmoset Monkeys Frontiers in pharmacology 8:395 doi:10.3389/fphar.2017.00395
Mitchell D (1976) Experiments on neophobia in wild and laboratory rats: a reevaluation. J Comp Physiol Psych 90: 190–197.D.
Muller L, Fritzsche P, Weinert D (2015) Novel object recognition of Djungarian hamsters depends on circadian time and rhythmic phenotype Chronobiology international 32:458-467 doi:10.3109/07420528.2014.992526
Nielsen KJ, Rainer G (2007) Object recognition: similar visual strategies of birds and mammals Current biology : CB 17:R174-176 doi:10.1016/j.cub.2007.01.014
Norris RW, Zhou K, Zhou C, Yang G, William Kilpatrick C, Honeycutt RL (2004) The phylogenetic position of the zokors (Myospalacinae) and comments on the families of muroids (Rodentia) Molecular phylogenetics and evolution 31:972-978 doi:http://dx.doi.org/10.1016/j.ympev.2003.10.020
Park HJ, Lee S, Jung JW, Kim BC, Ryu JH, Kim DH (2015) Glucocorticoid- and long-term stress-induced aberrant synaptic plasticity are mediated by activation of the glucocorticoid receptor Archives of pharmacal research 38:1204-1212 doi:10.1007/s12272-015-0548-0
Schleich CE (2010) Ontogeny of spatial working memory in the subterranean rodent ctenomys talarum Dev Psychobiol 52:592-597 doi:10.1002/dev.20466
Shao Y, Li JX, Ge RL, Zhong L, Irwin DM, Murphy RW, Zhang YP (2015) Genetic adaptations of the plateau zokor in high-elevation burrows Scientific reports 5:17262 doi:10.1038/srep17262
Sondergaard LV, Ladewig J, Dagnaes-Hansen F, Herskin MS, Holm IE (2012) Object recognition as a measure of memory in 1-2 years old transgenic minipigs carrying the APPsw mutation for Alzheimer’s disease Transgenic research 21:1341-1348 doi:10.1007/s11248-012-9620-4
Stefanko DP, Barrett RM, Ly AR, Reolon GK, Wood MA (2009) Modulation of long-term memory for object recognition via HDAC inhibition Proceedings of the National Academy of Sciences 106:9447-9452 doi:10.1073/pnas.0903964106
Stryjek R, Modlińska K, Pisula W (2012) Species Specific Behavioural Patterns (Digging and Swimming) and Reaction to Novel Objects in Wild Type, Wistar, Sprague-Dawley and Brown Norway Rats PloS one 7:e40642 doi:10.1371/journal.pone.0040642
Sutcliffe JS, Marshall KM, Neill JC (2007) Influence of gender on working and spatial memory in the novel object recognition task in the rat Behavioural brain research 177:117-125 doi:10.1016/j.bbr.2006.10.029
Uchida N, Mainen ZF (2003) Speed and accuracy of olfactory discrimination in the rat Nature neuroscience 6:1224-1229 doi:10.1038/nn1142
Wang K et al. (2017) Effect of 1.8 GHz radiofrequency electromagnetic radiation on novel object associative recognition memory in mice Scientific reports 7:44521 doi:10.1038/srep44521
Wegner RE, Begall S, Burda H (2006) Light perception in ‘blind’ subterranean Zambian mole-rats Anim Behav 72:1021-1024 doi:http://dx.doi.org/10.1016/j.anbehav.2006.02.018
Winkler H, Leisler B. Exploration and curiosity in birds: functions and mechanisms. In: Adams NJ, Slotow RH, editors. Proc. 22 Int. Ornithol. Congr. 1999. p. 915–32. Durban
Zhang X et al. (2015) The Effect of Oxytocin on Social and Non-Social Behaviour and Striatal Protein Expression in C57BL/6N Mice PloS one 10:e0145638 doi:10.1371/journal.pone.0145638
Zhang Y (2007) The Biology and Ecology of Plateau Zokors (Eospalax fontanierii). In: Begall S, Burda H, Schleich CE (eds) Subterranean Rodents: News from Underground. Springer Berlin Heidelberg, Berlin, Heidelberg, pp 237-249. doi:10.1007/978-3-540-69276-8_17
Zhang, Y.M., Fan, N.C. & Zhou, W.Y. (1993) Study on the population ecology of the plateau zokors. Chinese Journal of Vector Biology and Control 4, 359-361.
Zhou, W.Y. & Dou, F.M. (1990) Studies on activity and home range of plateau zokor. Acta Theriologica Sinica 10, 31-39.
Zoccolan D (2015) Invariant visual object recognition and shape processing in rats Behavioural brain research 285:10-33 doi:10.1016/j.bbr.2014.12.053
List of Figure
Figure (1) Diagram of the 2 recognition memory tasks and the delays used. (A) Novel Object Recognition task after 10 min (Short) and 24 hrs (long) delay interval (B) Object-in-place task after 30 min delay interval.
Figure (2) Mean duration (±SE) of contact time to objects during familiarization (A) and testing (B) sessions after 10-min retention period. * Significant at p ≤0.05.
Figure (3) Mean duration (±SE) of contact time to objects during familiarization (A) and testing (B) sessions after 24-hrs retention period.
Figure (4) Mean duration (±SE) of contact time to objects before (A) and after changing locations (B) after 10-min retention period. Bars with different superscripts are significant at p ≤0.05.
Figure (5) Mean (±SE) preference for novelty and place discrimination indices during familiarization and test sessions. * Significant at p ≤0.05.
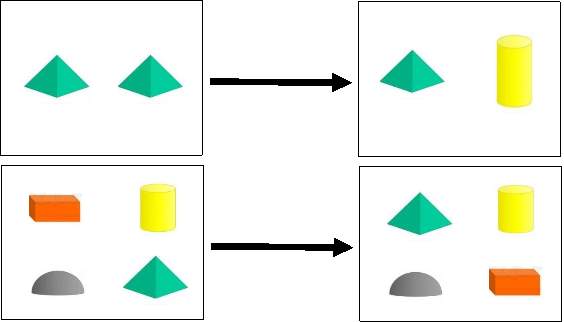
Figure (1)
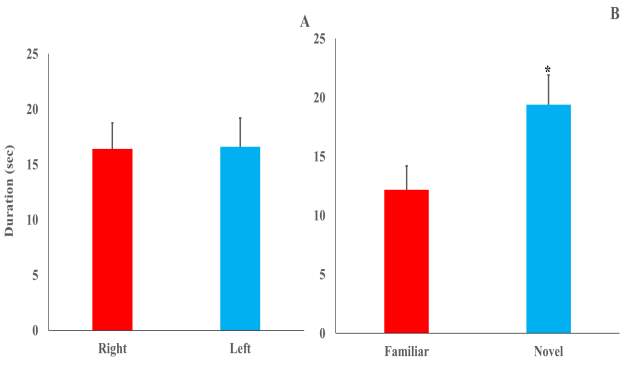
Figure (2)
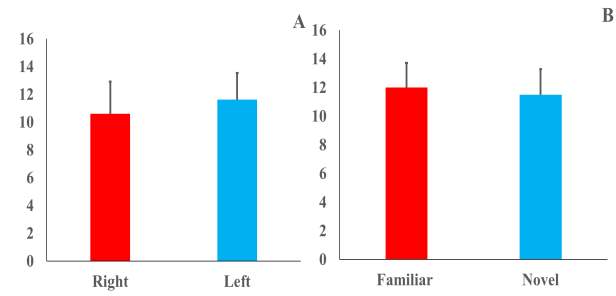
Figure (3)
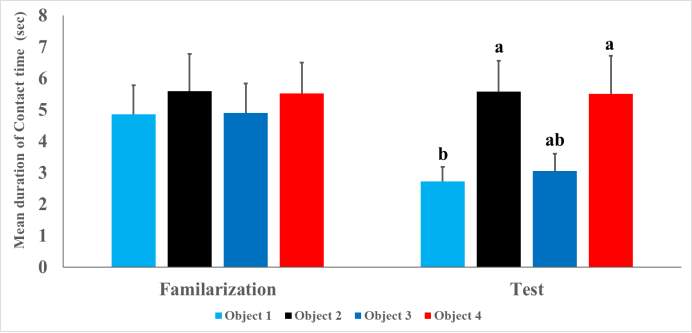
Figure (4)
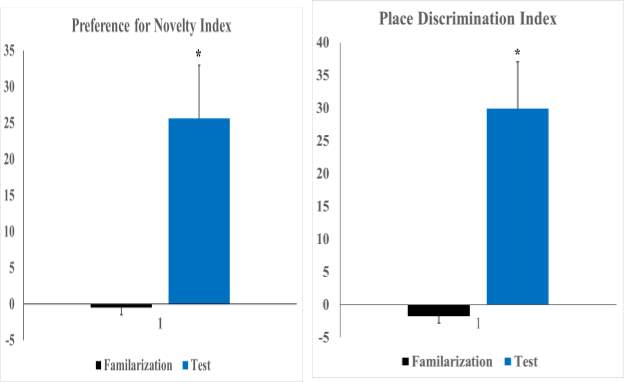
Figure (5)
Cite This Work
To export a reference to this article please select a referencing stye below:
Related Services
View all


Related Content
All TagsContent relating to: "Animal Sciences"
Animal science can be described as studying the biochemistry, physiology, and behaviour of animals that are under human control. Historically, animal science degrees were known as animal husbandry and focused on livestock. Studies now include companion animals such as cats and dogs.
Related Articles

DMCA / Removal Request
If you are the original writer of this dissertation and no longer wish to have your work published on the UKDiss.com website then please: